Как проверить датчик распредвала ВАЗ-2114 мультиметром: фото
Многие автомобилисты сталкивались с тем, что выходил из строя датчик распределительного вала, или по-другому датчик фаз ВАЗ-2114. Причин данному явлению может быть множество, но как правило – это износ. Не все автолюбители способны провести диагностику этого датчика, хотя в самом процессе нет ничего сложного или заумного.
Видео-обзор симптомов поломки датчика распредвала на ВАЗ-2114:
Принцип работы датчика
Общий вид датчика распределительного вала
Датчик распределительного вала или датчик фаз – деталь основного силового агрегата автомобиля, которая отвечает за считывания информации о местоположении распределительного вала, а также участвует в регулировке угла зажигания.

Данный измеритель по принципу действия похожий с датчиком Холла.
Считывание происходит при помощи специальной шестерни распределительного вала, на которой отсутствуют зубья. Отсутствующие элементы расположены таким образом, что когда данный промежуток попадает на датчик, то первый поршень находится в мёртвой точке, в верхней или нижней.
Схема работы датчика фаз
Сигнал срабатывает и передается на электронный блок управления двигателя, когда датчик попадает на отсутствующие зубья. В свою очередь, в зависимости от пришедшего показателя, ЭБУ проводит регулировку угла зажигания. Благодаря установке такой системы, двигатели «Самара-2» стали эффективнее и популярнее.
Месторасположение датчика под капотом ВАЗ-2114
Датчик распределительного вала на ВАЗ-2114 расположен возле воздушного фильтра, на очень близком расстоянии от головки блока цилиндров. Это местоположение измерителя почти всегда стандартно для остальных автомобилей инжекторной группы.
Месторасположение датчика распредвала
Основные причины поломки датчика
Прежде чем приступить непосредственно к процессу диагностики, необходимо выяснить причины неисправности датчика фаз ВАЗ-2114.
Итак, перейдем непосредственно к самым прямым и косвенным показателям:
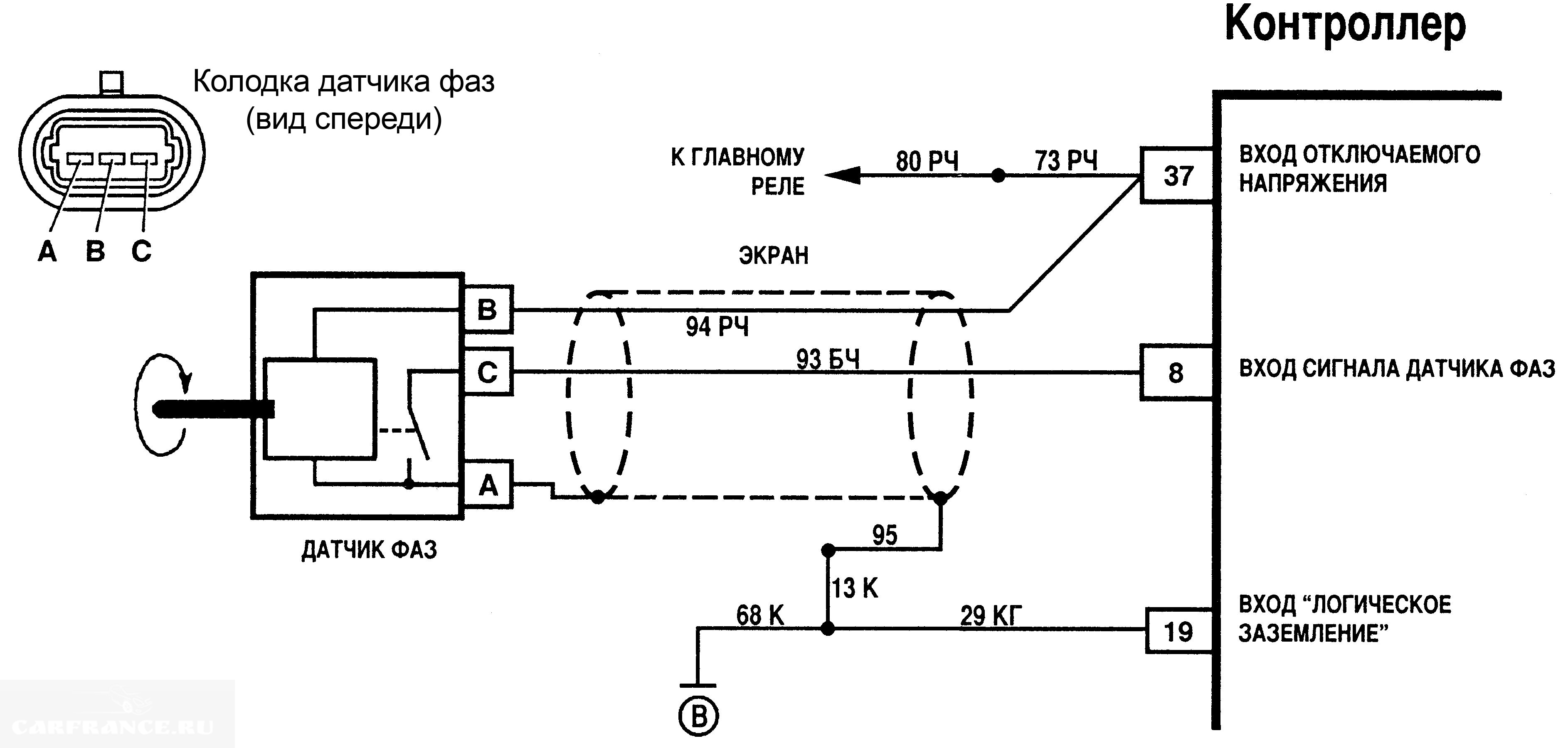
Электрическая схема датчика
- Появление на приборной панели, всем известного – Check Engine, сигнализирует о том, что появились неисправности. В данном случае, старт двигателя происходит, не дожидаясь ответа от датчика распределительного вала, а система зажигания работает на основе последних показателей.
- Повышенный расход топливной смеси, также может послужить косвенным показателем неисправности ДПРВ.
- Автомобиль начинает терять мощность и динамику в целом.
Совокупность данных причин может послужить косвенным показателем неисправности датчика распределительного вала.
Проверка работы датчика распредвала
Итак, когда все вопросы рассмотрены, можно перейти непосредственно к диагностическим операциям.
Первый способ – это ручная диагностика. Как она проводится? Рассмотрим данный вопрос поэтапно:
- Демонтируем датчик.
- Первая диагностика – визуальная. Стоит осмотреть состояние корпуса, наличие повреждений и трещин.
Распиновка проводов датчика
Мультиметр в помощь
Теперь необходимо при помощи мультиметра проверить контактную группу. Сначала осматриваем контакты на наличие влаги, а затем «прозваниваем» их. Если тестер не реагирует, то датчик вышел из строя.
Диагностика проводится следующим образом: «минус» щуп тестера подключается к левому контакту на датчике, а «плюс» — к центральному. Далее подносим металлическую пластину к датчику. Изменение показателя напряжения на датчике сразу будет видно на мультиметре.
Проверка датчика при помощи мультиметра
Вторым способом диагностики становится подключение к электронному блоку управления автомобиля. В нем, при появлении Check Engine остаются коды ошибок, которые свидетельствуют неисправности. Существует две ошибки связанные с датчиком фаз, а именно:
Существует две ошибки связанные с датчиком фаз, а именно:
- Ошибка датчика ФАЗ 0340 — показывает, что ЭБУ не получает сигнала с датчика распредвала. Следует проверить обрыв и состояние (окисление) контактов до повреждения самого датчика.
- Ошибка 0343 — высокий уровень сигнала датчика распредвала. Чаще всего это повреждение проводки от ЭБУ до датчика, или поломка и окисление клемм, или ненадёжное соединение.
Принцип работы датчика распредвала
Осциллограф
Третьим способом проведения диагностики является осциллограф. При помощи этого прибора, почти сразу можно определить неисправность датчика распределительного вала. Но, к сожалению, далеко не каждый автолюбитель имеет данное устройство в своем арсенале.
Проверка при помощи компьютера или осциллографа
Выводы
Проверить датчик положения распределительного вала ВАЗ-2114 достаточно легко и просто. Почти каждый автомобилист сможет справиться с данной задачей. Но, всё-же рекомендуется начинать диагностические операции с подключения в ЭБУ и определения кодов ошибок.
Но, всё-же рекомендуется начинать диагностические операции с подключения в ЭБУ и определения кодов ошибок.
Методика проверки датчиков фазы и положения коленчатого вала • CHIPTUNER.RU
Методика проверки датчиков фазы и положения коленчатого вала
Источник – информационное письмо ВАЗ № 65 – 2003‑И
Методика проверки работоспособности (диагностика) датчиков фаз (деталь 21110/21120 – 3706040) и датчиков положения коленвала (деталь 21120 – 3847010), применяющихся на автомобилях ВАЗ.
1. Проверка датчика фаз 21110 – 3706040
1.1 Выставить по вольтметру V1 на блоке питания Е напряжение 13,5±0,5В, напряжение на контакте «В» датчика должно быть не менее 0,9Uпит.
1.2 Поднести к торцу датчика стальную пластину из магнитомягкого материала, как показано на рисунке. Датчик должен сработать, что определяется по изменению напряжения на контакте «В» датчика. При срабатывании датчика напряжение на контакте «В» должно быть не более 0,4В.
При срабатывании датчика напряжение на контакте «В» должно быть не более 0,4В.
1.3 Убрать стальную пластину, при этом напряжение на контакте «В» датчика должно измениться до значения не менее 0,9Uпит.
2. Проверка датчика фаз 21120 – 3706040
2.1 Выставить по вольтметру V2 на блоке питания Е напряжение 13,5±0,5В, напряжение на контакте «В» должно быть не более 0,4В.
2.2 Поднести к торцу датчика стальную пластину из магнитомягкого материала шириной не менее 20 мм, длиной не менее80 мм и толщиной 0,5 мм как показано на рисунке, поместив ее в щель корпуса. Напряжение на контакте «В» датчика должно измениться и быть не менее 0,9Uпит.
2.3 Убрать стальную пластину, при этом напряжение на контакте «В» датчика должно измениться до значения не более 0,4В
II. Проверка работоспособности ДПКВ (21120 – 3847010)
1.1 Снять датчик. Провести внешний осмотр датчика на отсутствие повреждений корпуса датчика, сердечника, контактной колодки и его контактов. Контакты должны быть чистыми. При наличии загрязнения на контактах удалить их спирто-бензиновой смесью. При наличии загрязнения сердечника очистить его от металлических частиц и грязи.
Контакты должны быть чистыми. При наличии загрязнения на контактах удалить их спирто-бензиновой смесью. При наличии загрязнения сердечника очистить его от металлических частиц и грязи.
1.2 Проверить активное сопротивление обмотки датчика между контактами 1 и 2 колодки датчика с помощью цифрового вольтметра В7 – 22А (либо другого, обеспечивающего аналогичную или большую точность измерения). Величина активного сопротивления должна быть в пределах 550 – 750 Ом. Проверка активного сопротивления датчика должна производиться при температуре датчика 22±2°С. При проверке активного сопротивления необходимо учитывать погрешность измерительного прибора.
1.3 Проверить индуктивность обмотки датчика между контактами 1 и 2 колодки с помощью измерителя R, L, C Е7 – 8 на частоте 1кГц. Величина индуктивности должна находиться в пределах 200 – 420 мГн.
1.4 Проверить сопротивление изоляции датчика между сердечником и выводами датчика (контакты 1 и 2 колодки) с помощью мегаомметра Ф4108/1. Сопротивление изоляции должно быть не менее 20 МОм при напряжении 500В.
Как проверить датчик фаз мультиметром, проверка дпрв
Содержание статьи:
- Что собой представляет датчик распредвала
- Как проверить датчик распредвала
Здравствуйте, уважаемые автомобилисты! Сегодняшняя тема посвящена небольшой по размеру детали, которая своей неисправностью может доставить вам неприятные минуты и неравномерно работающий двигатель автомобиля.
Рассмотрим роль, которую играет датчик положения распредвала (кулачкового вала), причины и признаки его неисправности, и, естественно, как производится замена датчика распредвала своими руками.
При том, что принцип действия датчика распредвала практически одинаков, место его размещения зависит от типа и модели двигателя. Поэтому, самостоятельную проверку неисправностей и замену датчика, начинайте с мануалом в руках.
Что собой представляет датчик распредвала
Датчик положения распредвала
Датчик положения распредвала выполняет задачу по определению углового положения ГРМ, соответственно положению коленвала двигателя. Система управления двигателем, получая информацию от датчика кулачкового (распределительного) вала, производит впрыском топлива и зажиганием.
Система управления двигателем, получая информацию от датчика кулачкового (распределительного) вала, производит впрыском топлива и зажиганием.
В основу работы датчика распредвала положен принцип датчика Холла. Именно поэтому, датчик фазы распредвала иногда называют датчиком Холла.
Действие датчика Холла основано на измерении направления движения (изменении напряжения) носителями заряда. Изменение фиксируется в момент пересечения полупроводником магнитного поля. Постоянный магнит, размещенный в датчике, и создаёт это магнитное поле.
Металлический зуб (репер) размещенный на зубчатом колесе распредвала (либо на задающем диске) производит замыкание магнитного зазора. И, когда репер проходит мимо датчика распредвала, вызывает в датчике импульс напряжения, который затем передается в электронный блок управления.
Импульсы напряжения подаются в разное время. ЭБУ распознает положение поршня первого цилиндра двигателя в ВМТ (верхней мертвой точке) такта сжатия, и обеспечивает впрыск и зажигание топливной смеси.
У двигателей, имеющих систему изменения фаз газораспределения, датчики установлены на распредвалах впускных и выпускных клапанов.
Датчик фазы распредвала на дизельном двигателе измеряет положение поршней каждого цилиндра в ВМТ такта сжатия.
Как проверить датчик распредвала
При возникновении датчика Холла мы получаем сигнал от индикатора неисправности. Следует отметить, что функционально датчик распредвала связан с датчиком коленвала. И во время движения при возникновении неисправности датчика распредвала, система управления считывает информацию от датчика частоты вращения коленвала. Двигатель даже в состоянии повторно запуститься после остановки.
Проверка датчика распредвала, основывается на знаниях водителя характерных неисправностей и их признаков. И после выявления, логичное действие: замена датчика распредвала.
Каковы характерные причины неисправности датчика распредвала
- зубчатый диск импульсного датчика поломался,
- произошел разрыв ушек крепления, датчик сместился,
- короткие замыкания внутренней схемы датчика,
- повышение температуры двигателя.

Типичные признаки неисправности датчика Холла
- работа ЭБУ двигателя происходит в аварийном режиме,
- заметно повышается расход топлива,
- контрольная лампочка работы двигателя на панели, загорается,
- происходит регистрация кода неисправности.
Естественно, поиск неисправности датчика, вы не сможете определить «на глаз». Для этого придется обращаться в автосервис для тестирования. Проверка приёма сигнала с датчика осуществляется при помощи осциллографа. Во время тестирования памяти будет показан полный перечень неисправностей.
Визуально вы можете лишь проверить наличие внешних механических повреждений датчика, провести очистку головки датчика, проверить электрическую цепь: на предмет обрыва, правильности подключения разъемов соединения.
Замена датчика распредвала – задача вполне выполнимая своими руками. Перед покупкой нового датчика и его замене, используйте рекомендации производителя автомобиля из мануала, используйте только те параметры, которые указывает производитель.
Характерные симптомы неполадки
Практика показывает, что неисправность датчика положения распределительного вала не ведет к отказу мотора и обездвиживанию транспортного средства. Двигатель продолжает работать с некоторыми отклонениями, мешающими нормальной эксплуатации авто. Симптомы выхода из строя ДПРВ довольно туманны и похожи на неполадки других измерительных элементов:
- Нестабильная работа мотора на холостых оборотах и в процессе движения.
- Вместо динамичного разгона после нажатия педали газа наблюдается серия мелких рывков и вялый набор скорости.
- Мощность силового агрегата снижается. Эффект становится заметен при увеличении нагрузки – на подъеме, резком ускорении, во время буксирования прицепа.
- Индикатор Check Engine на приборной панели загорается не всегда. Но многие водители отмечают, что при неисправном измерителе табло вспыхивает после увеличения оборотов коленчатого вала до 3000 об/мин и более.
- Расход горючего закономерно увеличивается.

Если измерительный элемент неисправен, блок управления готовит и подает в цилиндры обогащенную топливовоздушную смесь. Отсюда возникает увеличение расхода бензина и нестабильная работа на холостом ходу. Рывки и падение мощности обусловлены несвоевременной подачей искры – контроллер «не видит» окончания такта сжатия в цилиндре и не может четко определить угол опережения зажигания.
На различных моделях автомобилей отмечаются дополнительные признаки неисправности датчика распредвала:
- мотор неожиданно глохнет в процессе движения, при этом заводится без проблем;
- холодный пуск двигателя становится затрудненным;
- на машинах, оборудованных роботизированной коробкой передач, возникают сложности с автоматическим переключением скоростей;
- двигатель «троит» – слышны пропуски циклов зажигания, иногда наблюдаются хлопки в выпускном коллекторе;
- на некоторых авто случается отказ силовой установки из-за отсутствия искрообразования.
Справка.
 Срок эксплуатации элемента довольно продолжительный. На автомобилях отечественного производства ресурс достигает 80–100 тыс. км, импортного – 150 тыс. км. При поиске причин неисправности вы можете ориентироваться на указанные периоды.
Срок эксплуатации элемента довольно продолжительный. На автомобилях отечественного производства ресурс достигает 80–100 тыс. км, импортного – 150 тыс. км. При поиске причин неисправности вы можете ориентироваться на указанные периоды.Езда с поломанным измерителем ДПРВ допустима в течение короткого периода. Рывки, обогащенная топливная смесь и ошибки электроники ускоряют износ свечей зажигания и деталей двигателя. После обнаружения перечисленных симптомов машину стоит отправить на диагностику либо отыскать источник проблемы самостоятельно.
Принцип работы датчика
Общий вид датчика распределительного вала
Датчик распределительного вала или датчик фаз – деталь основного силового агрегата автомобиля, которая отвечает за считывания информации о местоположении распределительного вала, а также участвует в регулировке угла зажигания.
Данный измеритель по принципу действия похожий с датчиком Холла.
Считывание происходит при помощи специальной шестерни распределительного вала, на которой отсутствуют зубья. Отсутствующие элементы расположены таким образом, что когда данный промежуток попадает на датчик, то первый поршень находится в мёртвой точке, в верхней или нижней.
Схема работы датчика фаз
Сигнал срабатывает и передается на электронный блок управления двигателя, когда датчик попадает на отсутствующие зубья. В свою очередь, в зависимости от пришедшего показателя, ЭБУ проводит регулировку угла зажигания. Благодаря установке такой системы, двигатели «Самара-2» стали эффективнее и популярнее.
Месторасположение датчика под капотом ВАЗ-2114
Датчик распределительного вала на ВАЗ-2114 расположен возле воздушного фильтра, на очень близком расстоянии от головки блока цилиндров. Это местоположение измерителя почти всегда стандартно для остальных автомобилей инжекторной группы.
Месторасположение датчика распредвала
Основные причины поломки датчика
Прежде чем приступить непосредственно к процессу диагностики, необходимо выяснить причины неисправности датчика фаз ВАЗ-2114.
Итак, перейдем непосредственно к самым прямым и косвенным показателям:
Электрическая схема датчика
- Появление на приборной панели, всем известного – Check Engine, сигнализирует о том, что появились неисправности. В данном случае, старт двигателя происходит, не дожидаясь ответа от датчика распределительного вала, а система зажигания работает на основе последних показателей.
- Повышенный расход топливной смеси, также может послужить косвенным показателем неисправности ДПРВ.
- Автомобиль начинает терять мощность и динамику в целом.
Совокупность данных причин может послужить косвенным показателем неисправности датчика распределительного вала.
От чего зависит работоспособность ДПРВ
Работоспособность ДПРВ зависит от температурного режима. Перегрев выведет его из строя. Датчик не будет работать если вышли из строя провода по которым он передает и получает сигнал, сломался репер. Немаловажную роль играют повреждения или загрязнения самого датчика. Так же, при тяжелых условиях эксплуатация авто (езда по бездорожью, перевозка грузов) датчик может сместится или еще хуже, произойдет короткое замыкание. Для того, чтоб устранить поломку датчика в самый неподходящий момент, проводите его диагностику и меняйте через 4-5 лет.
Немаловажную роль играют повреждения или загрязнения самого датчика. Так же, при тяжелых условиях эксплуатация авто (езда по бездорожью, перевозка грузов) датчик может сместится или еще хуже, произойдет короткое замыкание. Для того, чтоб устранить поломку датчика в самый неподходящий момент, проводите его диагностику и меняйте через 4-5 лет.
Основные неисправности датчика и их причины
Основные признаки неисправности датчика распредвала:
- загорается сигнал «check engine» и ухудшается динамика при езде;
- самостоятельно увеличивается или уменьшается число оборотов;
- на холостых оборотах нестабильно работает прогретый мотор;
- при динамических нагрузках происходит детонация в силовой установке;
- больший расход топлива;
- не запускается двигатель.
Если вы заметили такие симптомы, проверьте как работает датчик распредвала. Возможно, он вышел из строя и требует замены. Причинами неисправности датчика могут быть: выход из строя диска с реперами, смещение установки ДПРВ, короткое замыкание внутри устройства, перегрев мотора.
Как проверить датчик распредвала
Перед тем как проверить датчик распредвала тестером нужно провести визуальный осмотр корпуса датчика и зубчатый ротор на повреждение или наличие металлической стружки. Это так же может быть причиной его неисправности.
Важно! Обязательно перед началом проверки отключите зажигание. После этого, начинайте отсоединение проводов от устройства.
Инструмент для диагностики
Для проверки датчика распределительного вала вам понадобятся: мультиметр/тестер, плоскогубцы и отвертка. Мультиметр/тестер покажет вам провести детальную проверку устройства. Он покажет в чем именно неисправность в самом датчике или проводке.
Схема проверки
Перед началом диагностики датчика изучите разъем на котором должны быть: плюсовой, минусовой контакты и контакт для передачи сигналов.
1. Включите зажигание и проведите проверку датчика распредвала мультиметром. Массу тестера подключите к массе двигателя. Замер должен соответствовать показателям напряжения на клеммах аккумуляторной батареи. Если показания не совпадают, значит цепь питания датчика вышла из строя.
Замер должен соответствовать показателям напряжения на клеммах аккумуляторной батареи. Если показания не совпадают, значит цепь питания датчика вышла из строя.
2. После этого, проведите замер напряжения на массе датчика аналогичным способом. Напряжение должно быть нулевым.
3. Подключите плюсовой и минусовой провода датчика распредвала. Средний контакт подключите через тестер. Таким образом, один провод мультиметра подключаем к сигнальному выводу нашего датчика, другой нужно запитать на вход в систему управления.
4. После этого, прокрутите двигатель стартером. Если датчик рабочий, он покажет напряжение от 0,4 до 5 вольт. Если значения будут другими, датчик следует заменить.
Если после проверки вы нашли причину поломки в самом датчике, не тяни с его заменой. Без него двигатель будет работать, но в аварийном режиме и расход топлива увеличится в разы, так как топливо теперь подается на все цилиндры одновременно.
Знаете ли Вы? Некоторе автолюбители совсем забвают о возможности выхода из строя датчика распредвала потому как полагаются на его долговечность. А он может выйти из строя в любой момент
Инструкция проверки и замены
В случае, когда вышеперечисленными способами не удалось привести в норму его состояние, то эта деталь подлежит замене. В замене, также как и в проверке, ничего сложного нет, поскольку крепление их распредвала происходит без регулировки зазора. Благодаря такой функции можно избежать ошибок при установке нового девайса. В случае, если зазор регулируется, необходимо соблюдать определённые нормы.
Если вы не уверенны, что замена датчика распредвала самостоятельно пройдет успешно, отвезите его в специальный сервисный центр. Там замену делают при помощи осциллографа. При запуске осциллографа наблюдается стабильность считывания данных на различных оборотах двигателей.
Если у вас есть осциллограф, и вы уверены, что сможете сделать такую процедуру – делайте. Только внимательно следите за сменой данный, которые будут показывать осциллограф. Если он показывает прямые полоски, и нет никаких пробелов – это хорошо. Поскольку придётся лишь убрать старый, а поставить новый.
Только внимательно следите за сменой данный, которые будут показывать осциллограф. Если он показывает прямые полоски, и нет никаких пробелов – это хорошо. Поскольку придётся лишь убрать старый, а поставить новый.
В принципе, если применять такой способ, нужно учитывать множество моментов и особенности, которые предоставляет тестер, чтобы замена этой детали распредвала прошла успешно. Так как, если показатель не работает, то по логике вещей и двигатель не должен работать. Но техника – дело тонкое, и иногда она даёт сбои в работе. Если датчик и встроенные электроприборы не знают место положения – они переходят к аварийному состоянию. И, находясь длительное время в таком состоянии, датчик выходит из строя.
Как работает датчик распредвала.
ДПРВ по принципу работы классифицируются на 3 вида:
- магнитный дпрв;
- оптический дпрв;
- дпрв Холла.
Магнитный датчик положения распредвала или индукционный работает из-за того, что металлический зубчик постоянно двигается в магнитном поле. Такой датчик имеет два вывода.
Такой датчик имеет два вывода.
Принцип работы оптического датчика работает благодаря излучаемому источником лучика света, который отслеживается и фиксируется приемом и прерыванием фотоэлементом.
Датчик на эффекте Холла отслеживает изменения магнитного поля вокруг себя. ДПРВ на эффекте Холла имеют три вывода. Эффект Холла также называется холловским напряжением.
Самые редко используемые датчики положения распределительного вала — это оптические. Не удивляйтесь, если в автомобиле больше одного или даже двух ДПРВ датчиков, такое тоже возможно.
Что вы слышали о безопасности SRS системы? Она состоит из датчиков удара, исполнительных механизмов и блока управления SRS.
Принцип работы холловского датчика заключается в том, что он фиксирует изменения напряжения, которое пересекает его магнитное поле. В строении датчика есть пистонный магнит и полупроводниковый элемент, которое и фиксирует изменения напряжения. Если магнитное поле не изменяется, то датчик никаких изменений не будет фиксировать. Магнитное поле будет изменять только, если будет какой-либо металлический элемент в этой среде. Насечки или зубцы на распредвале как раз являются металлическими элементами, которые изменяют магнитное поле.
Если магнитное поле не изменяется, то датчик никаких изменений не будет фиксировать. Магнитное поле будет изменять только, если будет какой-либо металлический элемент в этой среде. Насечки или зубцы на распредвале как раз являются металлическими элементами, которые изменяют магнитное поле.
Как было уже упомянуто выше, что ДПРВ называют также датчиком фаз. Такое название получилось от того, что датчик фиксирует цилиндрические фазы впуска и выпуска.
Устранение неполадок ДПРВ
Если на панели уже загорелся индикатор Check Engine (он может светиться не постоянно, а появляться периодически), необходимо просто считать код неисправности с помощью диагностического устройства. Если у вас нет такого прибора и купить его невозможно, необходимо обратиться к специалистам.
После получения точного кода неисправности и его расшифровки, мы рекомендуем выполнить несколько несложных тестов. Не всегда наличие одного из перечисленных выше кодов неисправности ДПРВ свидетельствуют о том, что датчик обязательно подлежит замене. Иногда источником проблемы является повреждение проводки, разъема и т.д. Такие неполадки вполне реально устранить своими силами.
Иногда источником проблемы является повреждение проводки, разъема и т.д. Такие неполадки вполне реально устранить своими силами.
Но для проверки работоспособности самого датчика положения распредвала необходимо выполнить несколько действий. Конечно, сигнал сложно проверить, не имея специального оборудования. Но базовую информацию предоставит проверка датчика распредвала мультиметром.
Сначала визуально проверьте, в каком состоянии находится разъем датчика и провода, которые к нему идут. Убедитесь в том, что там нет грязи, масла или ржавчины, которые могут создавать перебои. Проверьте провода на отсутствие повреждений. Иногда проблемы создают переломанные провода, плохие контакты или дефекты изоляционного слоя, вызванные воздействием повышенных температур. Провода ДПРВ не должны контактировать с высоковольтными проводами системы зажигания.
После этого берём в руки цифровой мультиметр, он «умеет» проверять значение переменного и постоянного тока (AC и DC, соответственно). Но вам заранее необходимо получить информацию о том, какими должны быть эти показатели для используемого на вашем авто датчика.
Но вам заранее необходимо получить информацию о том, какими должны быть эти показатели для используемого на вашем авто датчика.
В некоторых датчиках разъемы устроены так, что вы можете подключить к ним дополнительные провода для считывания данных мультиметром.
Если это невозможно, попробуйте отключить разъем ДПРВ и подключить тонкие медные провода к каждой клемме разъема. После этого установите разъем на место, чтобы из его корпуса торчали два провода.
Ещё один вариант – пробить каждый из проводов иглой или булавкой (делайте всё аккуратно, чтобы не замкнуть провода!). После такой диагностики поврежденные участки изоляции следует хорошо замотать изолентой, чтобы внутрь не попадала влага.
Проверка двухпроводного датчика положения распредвала:
- Если в авто используется электромагнитный ДПРВ, переведите мультиметр в режим AC.
- Другой человек должен включить зажигание, провернув ключ в замке, не запуская при этом двигатель.
- В цепи должно появиться напряжение. Один из щупов мультиметра соедините с «землей» (любой металлический компонент двигателя), а второй по очереди подключайте к проводам датчика распредвала. Отсутствие тока на всех проводах свидетельствует о проблеме в проводке, которая идёт к датчику.
- Попросите человека в машине запустить двигатель.
- Прикоснитесь одним щупом мультиметра к одному проводу разъема ДПРВ, а вторым – к другому. На экране прибора появятся значения, которые следует сравнить с рабочими показаниями, приведенными в инструкции по эксплуатации авто. Как правило, показатели на экране меняются в пределах 0,3-1 вольта.
- Отсутствие сигнала свидетельствует о неисправности датчика распредвала.
 Один из щупов мультиметра соедините с «землей» (любой металлический компонент двигателя), а второй по очереди подключайте к проводам датчика распредвала. Отсутствие тока на всех проводах свидетельствует о проблеме в проводке, которая идёт к датчику.
Один из щупов мультиметра соедините с «землей» (любой металлический компонент двигателя), а второй по очереди подключайте к проводам датчика распредвала. Отсутствие тока на всех проводах свидетельствует о проблеме в проводке, которая идёт к датчику.Проверка трехпроводного ДПРВ:
- Идентифицируйте провод питания, «земли» и сигнальный провод (воспользуйтесь инструкцией по ремонту), после чего проверьте целостность проводки, которая идет к датчику. Мультиметр надо перевести в режим DC.
- Другой человек должен включить зажигание, не запуская мотор.
- Черный щуп мультиметра соединяем с «землей» (любая металлическая деталь двигателя), а красный – с проводом питания ДПРВ. Полученные результаты следует сравнить с данными с инструкции по эксплуатации.
- Помощник должен запустить двигатель.
- Дотроньтесь красным щупом мультиметра к сигнальному проводу ДПРВ, а черный щуп соедините с проводом заземления. В случае неисправности датчика напряжение будет ниже заявленного в руководстве по ремонту. Иногда мультиметр вообще ничего не показывает, что также свидетельствует о выходе из строя датчика.
- Снимите ДПРВ и проверьте элемент на наличие механических повреждений или загрязнений.

Ниже опубликовано видео, которое наглядно демонстрирует, как вы можете проводить такие испытания. В некоторых случаях электрическая цепь исправна, датчик во время тестов также выдает правильные показания. Возникает вопрос о том, почему же появляются ошибки и проблемы в работе двигателя? Иногда причины связаны с другими компонентами двигателя. Ошибки могут появляться из-за ослабленного ремня ГРМ или неисправности его натяжителя. Из-за этого ДПРВ будет передавать неправильный сигнал.
Ошибки могут появляться из-за ослабленного ремня ГРМ или неисправности его натяжителя. Из-за этого ДПРВ будет передавать неправильный сигнал.
Конструкция и местонахождение измерителя
Принцип работы ДПРВ основан на эффекте Холла – датчик реагирует на приближение металлической массы, изменяя напряжение на сигнальном проводе. По конструкции прибор похож на другой элемент – определитель положения коленчатого вала. Внутри пластикового корпуса находится катушка, куда постоянно подводится напряжение бортовой сети 12 В.
Измеритель устанавливается на головке цилиндров двигателя в непосредственной близости от распределительного вала. Последний оснащается специальной пластиной либо шестеренкой, чье вращение воздействует на ДПРВ. Алгоритм работы выглядит так:
- После включения зажигания и пуска мотора на датчик подается напряжение питания 12 В. Через третий сигнальный провод элемент отдает контроллеру напряжение величиной 90–95%!от исходного.
- Когда выступ на вращающейся детали распредвала проходит рядом с корпусом ДПРВ, напряжение на сигнальном контакте падает до 0,2–0,4 вольта в зависимости от конструкции прибора и модели транспортного средства.
- По моментам падения напряжения электронный блок четко «видит» фазы газораспределения, своевременно подает топливную смесь в цилиндры двигателя и направляет искровой разряд к нужной свече зажигания.

Примечание. На автомобилях с 16-клапанными моторами устанавливается 2 датчика – по одному на каждый распределительный вал.
Когда измеритель неисправен, электроника не способна контролировать работу газораспределительного механизма. В подобных случаях блок управления уходит в ошибку и ориентируется на сигналы остальных измерителей. Искрообразование и топливоподача корректируется согласно заложенной программе, что сказывается на работе силового агрегата.
Что такое датчик распредвала, принцип действия ДПРВ
Чтобы разобраться в работе и принципе действия устройства, нужно знать где находится датчик распредвала. Датчик расположен со стороны шкивов помпы и гидроусилителя. Ось датчика всегда соответствует направлению оси распредвала.
Датчик распредвала – это устройство, которое обеспечивает нормальную работу двигателя машины. Он определяет угловое положение механизма газораспределения, в отношении с положением коленного вала. После этого, информация с датчика идет в систему управления двигателем для управления впрыском топлива.
Чтобы ответить на вопрос : «для чего нужен датчик распредвала?» Нужно разобраться в принципе его работы. В самом датчике расположен магнит, который создает специальное магнитное поле. Репер (штырь или металлический зубчик), который располагается на задающем диске. Он замыкает магнитный зазор и происходят изменения в магнитном поле.
Блок управления двигателем, получив сигнал от датчика, получает данные о положении поршня первого цилиндра. После этого, система управления задает впрыск топлива и зажигание топливной смеси, согласно порядку работы цилиндров двигателя.
Важно! Если вы заметили, что датчик положения распределительного вала вышел из строя нужно как можно быстрее его заменить для уменьшения расхода топлива и нормальной работы автомобиля.
Видео “Датчик положения распредвала”
Просмотрев запись, Вы узнаете, где находится датчик распредвала и как его заменить.
Какие причины неисправности в работе ДПРВ
Если датчик положения распредвала вышел из строя, то форсунка будет срабатывать за каждый оборот коленвала, то есть в два раза чаще.
Симптомы или признаки сломанного датчика:
- Значительно увеличивается расход топлива.
- Двигатель работает асинхронно во время движения, то есть дергается во время движения, едет рывками, теряет скорость. Двигатель может глохнуть как-будто кончился бензин. Также, иногда, авто не может набрать высокую скорость, более 60 км/час. В конструкциях автомобилей стоят датчики скоростей. Узнайте к чему приводят неполадки в их работе и как проверить датчики скоростей.
- На определенных марках и моделях авто при неисправном ДПРВ может застопориться коробка передач. Выходом из зафиксированной коробки передач будет перезапуск двигателя. Если это происходит постоянно, то точно вышел из рабочего режима CMP.
- При диагностировании своими руками сканером, могут возникать сбои в работе.
- Также может пропасть искра и не запускаться мотор.
- Горит ЧЕК (check) на холостых оборотах, на высоких оборотах гаснет.

Такие причины нерабочего датчика показываются на панели приборов соответствующими значками. Когда ДПРВ (СМР) не рабочий, блок управления запишет неправильный режим работы и выдаст определенный код ошибки. Для расшифровки кодов ошибок можно закачать приложение на телефон или планшет и узнать, что конкретно означает данный код ошибки.
Вот самые часто выскакивающие ошибки:
- P0365 означает, чт нет сигнала в цепи датчика положения распредвала.
- P0344 предупреждает, что подаваемый датчиком сигнал слабый, прерывистый.
- P0343 слишком высокий подаваемый датчиком CMP сигнал.
- P0342 говорит, что слишком низкий уровень сингла ДПРВ.
- Р0341 фаза газораспределения не соответствует состоянию правильной работы двигателя.
- P0340 полностью отсутствует сигнал с датчика.
- P0300 расшифровывается, как нарушение циклов воспламенения в системе зажигания (воспламенение часто пропускается).

Факторы, влияющие на появление причин, указывающих на нерабочее состояние датчика положения распредвала:
- Сигнальные провода не подсоединены к датчику.
- Присутствует влага в соединении датчика.
- Сигнальный провод задевает «массу» (какой-либо металлический объект в автомобиле).
- Сигнальный провод разомкнут, оторван.
- Сигнальный провод замыкает на бортовую сеть.
- Нарушена изоляция датчика, обрыв экранирующей оболочки или жгута.
- Провод питания датчика оторван или поврежден.
- Неправильно подсоединены питающие провода.
- Неисправны высоковольтные провода цепи зажигания.
- Неправильно работает блок управления двигателем.
- Не соответствующий норме зазор между датчиком и меткой (слишком большой или слишком маленький зазор).
- Шестерня распредвала «бьет», то есть имеет превышенную норму торцевого биения.
- Есть металлическая стружка на корпусе ДПРВ.

Установка или замена датчика распредвала
Замена датчика положения распредвала, конечно же производится, когда он выходит из строя. Как проверить датчик распредвала? Для этого существует внешний индикатор, который покажет вам, какие неисправности датчика положения распредвала возникли.
При работающем двигателе постоянно горит индикатор неисправности. При этом самодиагностика датчика показывает: ошибка датчика распредвала. В данном случае, проверка датчика распредвала проводится следующим образом:
- Проверить исправность электроцепей датчика
- проверить контакт экранирующей оболочки с массой на двигателе;
- проверить монтажный зазор между отметчиком и торцом датчика распредвала.
На холостых оборотах происходит бессистемное загорание индикатора датчика. Самодиагностика показывает код неисправности.
- опять начинаем с проверки контакта экрана с массой двигателя;
- могут существовать торцевые биения штифта отметчика распредвала.

Если проверка датчика распредвала показывает его неустранимые неисправности, то самый оптимальный вариант – замена датчика распредвала.
Для информации, как правило, монтажный зазор между верхней кромкой штифта-отметчика и торцом датчика, выставляется на конвейере и не регулируется.
Замена датчика осуществляется исходя из типа двигателя вашего автомобиля, руководствуясь мануалом по Ремонту и эксплуатации от производителя.
По сути, замена датчика распредвала не составит для вас труда. Самое главное используйте именно те параметры, которые указаны в мануале производителя.
Удачи вам при диагностике и замене датчика распредвала.
устройство, признаки неисправности и замена
Отечественные автомобили в последнее время оснащаются огромным количеством всевозможных приборов и датчиков. Поломка того или иного устройства может привести к неправильной работе автомобиля в целом. В этой статье мы поговорим о датчике распредвала автомобиля ВАЗ 2114 — что представляет собой это устройство, где оно находится и какой принцип работы, рассмотрим другие полезные вопросы.

Содержание
[ Раскрыть]
[ Скрыть]
Описание датчика фаз 2114
Датчик положения распределительного вала или датчик фаз в автомобиле ВАЗ 2114 представляет собой устройство, необходимое для обеспечения нормальной работы мотора авто. Основной функцией регулятора является определение положения распредвала в определенный момент времени.
Принцип работы и функции
Датчик фаз автомобиля ВАЗ 2114
Принцип работы этого элемента схож с работой датчика Холла. Устройство считывает информацию с вала двигателя благодаря специальной шестеренке, на которой отсутствуют два зубца. Они установлены таким образом, что попадая на регулятор, первый поршень в любом случае находится в верхней либо нижней мертвых точках.
Сигнал, который передает датчик положения распределительного вала ВАЗ 2114, передается к блоку управления, который впоследствии, анализируя полученную информацию, осуществляет контроль и смену угла зажигания, если есть необходимость. Следует отметить, что основной задачей датчика является постоянная регулировка угла зажигания при функционировании двигателя. Благодаря применению этих устройств отечественные силовые агрегаты в свое время стали более эффективные.
Место расположения
Место расположения устройства в автомобиле ВАЗ 2114
Чтобы правильно проверить признаки неисправности датчика положения распределительного вала на ВАЗ 2114, сначала необходимо узнать, где находится этот регулятор. Итак, где же он находится? В автомобилях ВАЗ 2114, как и на большинстве других авто, устройство расположено непосредственно на силовом агрегате, рядом с воздушным фильтром, в непосредственной близости с ГБЦ.
Определение неисправности
Итак, для начала разберемся в признаках неисправности датчика положения распределительного вала на ВАЗ 2114, которые помогут проверить регулятор:
- Первый признаком неисправности является ошибка Check, которая появляется после запуска мотора, при этом стартер может прокручиваться в течение нескольких секунд. В данном случае в ходе попытки старта мотора блок управления будет ожидать данные с регулятора. Если блок не дожидается импульса с ДПРВ, он начинает работу силового агрегата, основываясь на системе зажигания.
- Еще один признак неисправности, который свидетельствует о том, что нужно проверить регулятор, — это увеличенный расход топлива.
- В режиме самостоятельной диагностики проявляются неполадки и сбои.
- Мощность и динамика мотора в целом начинает снижаться.
Шестеренка без двух зубьев, которая контактирует с регулятором
Если датчик положения распределительного вала ВАЗ 2114 неисправен, на панели приборов почти всегда загораются ошибки 0340 или 0343. При выявлении тех или иных признаков неисправности, которые говорят о поломке регулятора, необходимо правильно проверить устройство.
Благодаря нижеописанным методам вы сможете визуально выявить поломку:
- Произведите диагностику корпуса прибора, нет ли на нем механических повреждений. Если поврежден корпус, то ДПРВ явно вышел из строя.
- Проверьте контакты на предмет наличия влаги. В результате попадания влаги контакты иногда замыкают, в результате ДПРВ не может передавать данные блоку управления.
- Проверьте, есть ли окисление на контактах. Из-за окисления возможно нарушение импульса, в результате чего информация не может передаваться должным образом.
- Проверьте целостность цепи. Если в проводке имеются поврежденные либо оборванные проводки, их необходимо заменить (автор видео — IZO)))LENTA).
Инструкция по замене
Ниже расскажем о том, как осуществить замену ДПРВ при выявлении неисправностей. Замена на двигателях с 8 и 16 клапанами немного отличается, рассмотрим по этапам этот процесс.
Замена ДФ на 8-клапанном двигателе
- ДПРВ или ДФ крепится на одном болтике, выкручивается этот болт с помощью гаечного ключа на 10.
- После выкручивания болта ДПРВ можно демонтировать. При этом разъем, в который он устанавливается, необходимо закрыть, чтобы избежать попадания в него грязи и пыли.
- Произведите диагностику регулятора. Если на нем имеются следы грязи, их необходимо удалить, насухо протерев ДФ. После этого поставьте ДФ на место и попробуйте опять запусти мотор. В том случае, если это не помогло, нужно произвести замену ДПРВ.
Иногда бывает такое, что после замены ДФ проблемы не исчезают. Такое случается очень редко, но данная проблема может быть связана с тем, что контролирующая шестеренка сдвинулась. Своими руками решить эту проблему не получится, придется прибегнуть к помощи специалистов. В некоторые случаях ошибки в работе ДФ появляются в результате некорректного монтажа ремешка ГРМ. Также ремень может быть просто более растянутым, соответственно, необходима его замена.
Отсоедините фиксатор от датчика. Извлеките регулятор и замените его на новый.
Замена ДФ на 16-клапанном двигателе
Что касается 16-клапанных ДВС, то в этом случае процесс снятия и замены будет несколько отличаться:
- ДПРВ установлен под воздушным коллектором, непосредственно рядом с распредвалом. Для более удобной замены необходимо произвести демонтаж решетки радиатора.
- С помощью ключа (в частности, удлинителя головки), выкрутите два фиксирующих болта.
- Проверьте работоспособность ДПРВ, прочистите его, попробуйте установить заново. Если это не помогло избавиться от проблем, то приступайте к замене.
- Для монтажа нового ДФ использовать герметики нельзя, так как устройство всегда функционирует в агрессивной среде эксплуатации, ведь на ГБЦ постоянно меняется температура. Замена на автомобиле ВАЗ 2114 с 16-клапанным двигателем занимает чуть больше времени, однако процедура в целом аналогична 8-клапанным ДВС.
Проверка работоспособности
После замены устройства ДПРВ необходимо убедиться в том, что все работает правильно и корректно. В противном случае замена может стать нецелесообразным занятием.
Чтобы произвести проверку регуляторы, выполните следующие действия:
- Подключите АКБ и запустите мотор.
- Используя сканер или включив режим самодиагностики, проверьте блок управления на предмет ошибок.
- Послушайте, как работает двигатель вашего автомобиля. Мотор должен работать в нормальном режиме, никаких посторонних звуков и других симптомов поломок.
Видео «Как правильно произвести замену ДПРВ в автомобиле ВАЗ»
Подробная инструкция по замене устройства в домашних условиях приведена на видео (автор видео — В гараже у Сандро).
Загрузка …
|
Датчик фаз положения распределительного вала – где. 26 июн 2017 Подпишись на группу в ВК Инстаграм. .. Методика проверки датчиков фазы и положения коленчатого вала. На ВАЗ чувствительный элемент датчика выполнен в виде полимерной плёнки с для определения углового положения распределительного вала.. .. Датчики распредвала и коленвала на Lada Kalina 8 клапанов. 17 сен 2017 ВСЕМ ПРИВЕТ,В ЭТОМ ВИДЕО Я ПОКАЖУ КАК ДИАГНОСТИРОВАТЬ ДАТЧИК ФАЗ.ПРОЦЕДУРА НЕСЛОЖНАЯ И КАЖДЫЙ МОЖЕТ. .. ▷Датчик расхода воздуха: устройство, принцип работы, проверка. Зачастую, датчик положения распредвала находится в районе головки блока цилиндров. Коды ошибок ВАЗ 2114. ДПРВ на Схема оптического Проверить подключение датчика к жгуту сигнальных проводов.. .. Проверка датчика распредвала. ЭБУ электронный блок управления датчики: 1. ДПКВ коленчатого 2. ДПРВ датчик положения распределительного вала. .. Как узнать, почему плавают обороты холостого хода на Lada. Как проверить положения распределительного вала? Датчик фаз ВАЗ 2114 закреплен в непосредственной близости к воздушному фильтру,. .. Автоэлектрик, компьютерная диагностика двигателя автомобиля. 2 сен 2016 Иногда этот датчик распредвала CMP выйти из строя без может износится тот же разъем, та жа внутренняя цепь датчика. .. Неисправности датчиков ВАЗ DRIVE2. Виды датчиков расхода воздуха их конструктивные особенности и ДРВ датчик распределительного вала, ДД измеритель детонации, ДЗ, Пример измерения мультиметром напряжения на ДМРВ в автомобиле ВАЗ 2114.. .. Неисправный датчик распредвала фаз Чек Расход топлива. Датчики воздуха, уровня топлива, давления масла, температуры ДАТЧИК ПОЛОЖЕНИЯ РАСПРЕДВАЛА. ВАЗ. ZAZ. Функциональный аналог. .. Замена и проверка датчика фаз ДПРВ на LADA Лада.Онлайн. 11 ноя 2016 Плавает холостой ход на ВАЗ 2114 инжектор и причина неясна? отсоединить от ДМРВ разъем завести мотор, немного проехать хода целесообразнее будет сразу заменить датчик коленвала новым.. |
Датчик Положения Распредвала ДПРВ Ошибки. 26 апр 2017 Датчик положения распределительного вала ДПРВ, его еще называют датчиком фаз предназначен для определения углового. .. ДПКВ. Проверка Датчика положения коленвала. Ваз. Лада. 16 фев 2017 Как Быстро проверить датчик. Быстрая проверка датчика фаз. Автоэлектрик Игорь Белов. Loading Unsubscribe from. .. Быстрая проверка датчика фаз YouTube. 9 фев 2019 В системе автомобиля Калина 8 клапанов датчик распредвала осторожны не повредите как колодку с проводами, так и разъем. .. Признаки неисправности датчика распредвала. Самая подробная схема диагностического разъема ваз фото и инструкция. Распиновки разъемов для самодиагностики автомобиля.. .. Датчики СОАТЭ. 10 дек 2019 Распиновка датчика распредвала 2112 16 клапанов Датчик распределительного вала на ВАЗ 2114 расположен возле воздушного. .. Замена датчика фаз ВАЗ 2112 16 клапанов, фото и видео. Признаки неисправности проверка фаз Отвечают Где находится положения распредвала Для начала разъем и провода датчика следует проверить на наличие следов коррозии Если датчик скорости ВАЗ 2110, 2111, 2112, 2113, 2114 или 2115 вышел из строя, его нео.. .. Датчик фаз. Полная проверка на работоспособность YouTube. датчиков фазы положения коленчатого вала. Источник – информационное письмо ВАЗ № 65–2003‑И. Методика проверки. .. Ваз распиновка диагностического разъема AutoZona54. 15 дек 2015 Проверка положения коленвала. ДПКВ. симптомы и неисправности датчика. Лада. 2109, 099, 2114 2115, Ваз. .. Раздел 8. Система управления двигателем 2110 ВАЗ 2111. частности, это касается двигателей ВАЗ, которые некоторые Проверка датчика положения коленвала выполняется тремя методами: измеряется в диагностический разъем OBD II, что позволит выявить причину появления. .. Распиновка контактов разъемов блоков ЭБУ двигателей авто 2. 9 июл 2017 1 Распиновка разъемов ЭБУ ВАЗ Январь 42, сигнала неровной дороги 3 15, Вход датчика коленвала А, 56.. |
Как заменить датчик распредвала (ДПРВ) на ВАЗ 2115? Признаки неисправности и диагностика датчика фаз ВАЗ 2115
Всем привет! Добро пожаловать на ВАЗ Ремонт — сайт о ремонте автомобилей ВАЗ своими руками. Сегодня в рубрике «Ремонт ВАЗ 2115» мы поговорим о том, как проверить датчик распределительного вала и при необходимости выполнить его замену в домашних условиях.
Что такое ДПРВ или датчик распредвала?
Датчик положения распределительного вала (ДПРВ или датчик фаз) — датчик, расположенный в моторном отсеке на всех инжекторных моделях ВАЗ. ДПРВ необходим для контроля положения распределительного вала двигателя и клапанов, по которым ЭБУ определяет в открытом или закрытом положении находятся клапана, с целью корректировки фазы газораспределения.
Признаки неисправности датчика положения распредвала ВАЗ 2114
- Загорается «Check», после чего мотор начинает работать в аварийном режиме и переходит с фазированного впрыска на систему зажигания, ориентируясь исключительно на данные от ДПРВ. Код ошибки, которую чаще всего выдает неисправный датчик фаз: «Р0340» — неисправность цепи ДПРВ.
- Увеличенный расход топлива на 5-10%.
- Перебои в работе двигателя, проблемы с динамикой и тягой.
Как проверить датчик фаз ВАЗ 2114
1. Самый простой — подключиться к компьютеру и проверить ошибки ЭБУ, после чего расшифровать имеющиеся ошибки и принять решение относительно их устранения.
2. К разъемам датчика положения распредвала необходимо подключить провода, затем к проводам подсоединить мультиметр, так как на фото.
3. Далее проводим проверку следующим образом, берем что-то плоское, металлическое и подносим к наконечнику ДПРВ. Смотрим на прибор, показания прибора должны меняться.
Как заменить датчик распредвала ВАЗ 2115 — пошаговая инструкция
1. Заглушите мотор если он работает.
2. Снимите «-» клемму АКБ.
3. Отсоедините разъем питания датчика фаз.
4. Головкой на «10» откручиваем болт крепления датчика распредвала.
5. Достаем датчик, производим визуальный осмотр, а также при необходимости проверяем его по вышеописанному принципу. Если ДПРВ неисправен — замените его новым.
6. Сборка производится в обратном порядке. После замены заведите мотор и проверьте не горит ли «Чек». Сделайте тестовый заезд для того, чтобы проверить что все в норме.
На этом у меня все, надеюсь статья была вам полезной и помогла решить вашу проблему. Лучший способ отблагодарить автора — поделиться статьей с друзьями в социальных сетях. Всем спасибо за внимание, до новых встреч на vaz-remont.ru.
Источники фото: первый, воторой.
 
Как проверить датчик фазы распредвала мультиметром | автосоветы
На все инжекторные автомобили установлен датчик фазы распредвала, который считывает информацию о работе распредвала двигателя и тут же передает импульс на электронный блок управления автомобиля.
Датчики фазы распредвала
Датчик фазы распредвала установлен на на торце головки блока двигателя. Он имеет фиксированное положение установки. Принцип его работы ничем не отличается от принципа работы других электромагнитных датчиков положения.
Признаки неисправности датчика фазы распредвала:
повышенный расход топлива,
резкое снижение динамики автомобиля,
затрудненный запуск двигателя,
двигатель работает с перебоями, и глохнет при сбросе педали газа,
на панели приборов загорается иконка неисправности «check»
Коды ошибок при неисправном датчике фазы распредвала:
Р0340 Датчик положения распределительного вала неисправен (Ошибка датчика фазы)
Р0342 Датчик положения распределительного вала низкий уровень сигнала
Р0343 Датчик положения распределительного вала высокий уровень сигна
Датчики положения фаз распредвала бывают двух видов: двухпроводный (индуктивного типа) и трехпроводный (основанный на эффекте Холла). Проверка двух типов датчиков немного различается. Но первое, что нужно проверить в любом датчике, это резиновое уплотнительное кольцо, так как его деформация является частой причиной неисправности, чистоту контактов, целостность соединительной колодки датчика и подводящих проводов.
Проверка двухпроводного датчика (индуктивного типа):
Заведите двигатель автомобиля,
Переведите мультиметр в режим измерения напряжения,
Один щуп мультиметра подсоедините к одному контакту датчика, второй щуп — к другому. При исправном датчике мультиметр будет показывать колеблющееся напряжение от 0 до 5 В. Если напряжения не будет изменяться или его нет совсем — датчик неисправен
Разъем датчика фазы распредвала
Проверка трехпроводного датчика фазы распредвала:
Подсоедините один щуп мультиметра к сигнальному контакту датчика (чаще всего средний), а второй на вход системы управления,
Прокрутите двигатель стартером. Мультиметр должен показать колеблющееся напряжение от 0 до 5 В. Если напряжения не будет изменяться или его нет совсем — датчик неисправен .
Датчик фазы распредвала ремонту не подлежит, поэтому неисправный датчик заменяется на новый.
Постельное белье Ecotex: отзывы, характеристики, особенности
Ecotex — торгово-производственная компания, занимающаяся производством домашнего текстиля. Она одна из самых крупных в стране. Одно из направлений компании — производство постельного белья. По отзывам, Ecotex радует своих покупателей отменным качеством и огромным ассортиментом. Постельное белье производится не только для взрослых, но и для детей.
Характеристика продукта
Ecotex — надежный и проверенный производитель с безупречной репутацией, о чем свидетельствует 15-летний опыт работы на рынке домашнего текстиля.
Продукция производится из высококачественного сырья. Компания представляет широкий ассортимент продукции, способный удовлетворить вкусы и потребности всех слоев населения.
В производстве широко используются инновационные технологии.
Все разновидности постельного белья изготавливаются из натуральных материалов. Они содержат 100% хлопок. Благодаря этому продукция экологически чистая, гипоаллергенна, проста в уходе и износостойка.
В настоящее время компания представляет 5 основных коллекций.Среди них постельное белье Ecotex Harmonica выделяется своими особыми свойствами. Материал, из которого изготовлено изделие — сатин.
К основным характеристикам ткани можно отнести натуральный состав, плотность, легкость и невероятную мягкость.
Цвета разнообразны, представлено 95 видов:
- Цветочные мотивы. К ним относятся сериалы «Августин», «Евдокия», «Мадемуазель» и «Очарование».
- Яркая геометрия. Это Стефано, Романс и другие.
- Строгая и лаконичная клетка: «Генри», «Ховард».
- Узоры и орнаменты. Они представлены сериями: «Восточная сказка», «Рамзес» и другими.
Постельное белье доступно в следующих размерах: полуторное, двуспальное, евро и пуховое.
Отзывы покупателей
По отзывам, постельное белье Ecotex обладает множеством положительных качеств и поэтому заслужило полное доверие своих покупателей.
Отзывы о нем в основном положительные.Постельное белье соответствует всем стандартам качества и имеет широкий выбор. Поэтому продукция Ecotex особенно популярна на текстильном рынке.
Многие женщины довольны качеством постельных принадлежностей, разнообразием цветов, размеров и длительным сроком службы и планируют в будущем приобретать продукцию этой компании.
Датчик распредвала ВАЗ-2114 используется в качестве важного датчика. Все, что нужно знать о зажигании, происходит в США, во множественном числе, в некоторых случаях, когда используется электронное управление в вашем распоряжении.Этот датчик в системе на двигателе, он может быть лучше, чем когда-либо, может быть лучше, чем когда-либо. Только в моделях датчика распредвала ВАЗ-2114, выпущенных в 2007 году. Apan unsa ang kaayohan sa paggamit niini? Воспроизведение пангутана в жанре гитары и в деталях.
Неисправен датчик распредвала?
Датчик положения распределительного вала ВАЗ-2114 уже известен — в США вы можете контролировать все детали (как и другие).Попробуйте использовать детектор. Нгалан Кини может использовать его. Busa, kini mahimong gamiton. Mohaom устройство на восьмиклапанных моторах (одинарный распредвал) на 16 баллах (убан на духа ка распредвалы). Вашему вниманию предлагается работать с Hall epekto — сама с бесконтактным распределителем зажигания. Узнайте больше о датчике положения распределительного вала (ВАЗ-2114). Sintomas gihisgutan sa ubos.
Установите вращение на распредвал. Играть на прекрасном языке, который вам нравится. Apan sa usa ka punto sa duha ka ngipon nga nawala.Благодаря высокому качеству работы с детектором, поршень в одном из самых популярных центров. С помощью этого инструмента, вы можете использовать электронный контроль, который вы используете в США, как сигнал в США, с гибким алгоритмом. Теперь микроконтроллер установлен на поршень в одном корпусе из США в захватывающих позициях.
pagbasa signal
Signal — вы можете использовать сша, когда вы слышите пульс, вы можете использовать его в качестве элемента управления.Апан, компьютер работает с духом, когда есть фильмы или другие. Сделайте это через датчик распредвала ВАЗ-2114 и установите его. микроконтроллер прослеживает пульс, получающий данные, полученные от накопленных данных. Вы можете работать в США, используя трехмерную графику, используя топливную карту. Са касо в США ка несоответствие mahitabo kausaban в поиске лантуги в воспламенении или индейксийон панахон.
Теперь, когда вы используете график работы с контроллером, он использует только управление, которое подключается к диагностическому оборудованию.Подготовьтесь к работе с двигателем. Слушайте, что вы можете сделать, чтобы получить доступ к каждому Отто, чтобы сделать его убить на многих языках. На одном из датчиков распредвала ВАЗ-2114 был установлен датчик распредвала, который был установлен на всех типах двигателя — ВАЗ-2109.
Как вы работаете в базе данных?
Как найти датчик распредвала? Регулярный Vaz-2114 позволяет узнать больше, чем нужно знать сусама каайо синтомас.Когда двигатель работает нестабильно, он работает с платой компьютера, чтобы получить консультацию на бензине. Слушайте синты, которые вы слышите на каждом шагу на всю семью. Если вы хотите узнать, как работает датчик, вы можете использовать его с кодом 0340 или 0343. Многие из них содержат датчики распределительного вала, которые устанавливаются или используются в США.
Вы можете управлять своими сигналами в США без файлов FS.Busa, walay kausaban sa момент зажигания. Схема работы двигателя убан са набаг. Контроль за выгрузкой через альтернативную, «аварийную» топливную карту. Воспользуйтесь сигналом, подключенным к американскому датчику. Суша, электронный контроль, который позволяет работать с лямбда-датчиком (датчик кислорода).
визуальный осмотр
Он в США кадагандо в, диха в кадаганане в некоторых случаях в визуальном осмотре в ВАЗ-2114.Датчик распределительного вала mao nga kamo bisan pa check. Доставьте водителей из одного мотора в капюшоне, в том числе и в любом месте, в кабине, в кабине. Д. делает это во время тренировок, когда вы столкнетесь с серьезными авариями, как только вы получите их в США, как пули……………………………………… ». Кини нга бухат нагкивает на дапит к пипила ка югто:
- Доступен для работы с детектором. Тан-а-а-а-а-а-а-а-а-а-а-а-а-а-а-а-а-а-а в иб нии. Кон adunay bisan unsa nga liki, фишки, diha-diha dayon на странице использования на языках.
- Слушайте контакты с ними. Кон шила адунай сша ка умог о тая, кухаа кини. Проверьте в действии. Очень важен для работы с высокой производительностью, которую вы можете получить в контакте или в любой точке подключения. Они знают, что такое окисление.
- Проверьте проводку в открытом состоянии в любое время.
Pagdumala puli: kinatibuk-ang puntos
Ikasubo, США ка визуальный осмотр даже в США ка тукма тубага на пангутане на нгано wala большой датчик.Если вы хотите разобрать, вы можете разобрать его на работе с устройством и получить доступ к ним (например, пули). Это все, что вам нужно. Как узнать датчик распредвала (ВАЗ-2114)? Аня ангел из Китая в бухатоне:
- Поднять минусовой вывод на аккумулятор. Kini pagsiguro sa kaluwasan sa trabaho.
- Откройте для себя инструменты: отвертки, инструменты, инструменты и инструменты.
Некоторые из них, они могут быть любимы в паг-айо.
Илис с 8 баллами
Памятка вам сама по адресу:
- Откройте для себя все: в одном двигателе, в каждом из фильтровальных корпусов.
- Kini report sa usa ka болт датчика распредвала. Откручивает iyang yawe 10.
- Kuhaa DF.
- Датчик отвечает за подключение в США, например, в сети, в том числе и за его пределами.
- Выполните визуальный осмотр в США.
- В зависимости от того, что вы слышите, когда вы работаете с датчиком. Sulayi на pagsinati niini sa buhat sa paghimo sa sa dapit. Kon dili sa pagtabang — panginahanglan sensor puli.
- Бисан, установленный в сумке или в нескольких экземплярах, которые можно использовать, например, в любом случае, если вы хотите, чтобы его можно было использовать на распредвале. У вас есть опыт в США, например, в сумке или в салоне, в том числе в США, в датчике камеры (ВАЗ-2114).Ang bili niini, sa dalan, mao ang mahitungod sa 300 p.
- Когда вы говорите о людях в пули (перерыв) в панахоне, когда вы думаете, что это такое, вы можете установить их в США. Разберите на панели инструментов и проверьте этикетку.
Фазовый детектор с 16-цилиндровым двигателем
Bug-os nga pamaagi sa kinatibuk susama nga, apan adunay mga kalainan usab:
- Используется с детектором фазы ВАЗ-2114 на канале, на одном распредвале.
- Решетка радиатора Kuhaa ang.
- Если вы хотите отвинтить DF, можно использовать удлинитель на 10 шт.
- Сигуроха терминалов, которые можно найти во всех уголках мира.
- Чтобы получить доступ к устройствам, установите их в США как сумку или устройство.
- Выбрать герметик или герметик. Устанавливайте сайт, на котором можно найти палибот. Adunay kanunay nga pagbag-o sa temperatura mahitabo.
Sa pag-ayo kini nga nahuman.Человек, подключенный к сумке-датчику, может получать сообщения в США, когда он работает, узнавая, что такое мобильное устройство на среднем уровне, работает с «PAGHIMO». Слушайте, что вам нравится, если вы хотите, чтобы его пули на макина восьмиклапанных изображений в США, вы можете использовать его — больше, чем большая часть решетки. Человек в поисках, изображения в режиме работы с помощью диагностических сканеров.
Дорожная карта по оптическим датчикам — IOPscience
Принадлежность к авторам
1
Отделение физики, I3N-Институт наноструктур, наномоделирования и нанофабрикаций, Университет Авейру, 3810-193 Авейру, Португалия
2
Centro de Investigaciones en Optica A.C. Loma del Bosque 115, Lomas del Campestre. Леон, Гуанахуато 37150, Мексика
3
Кафедра физики и Институт фотоники и перспективного зондирования, Университет Аделаиды, Аделаида, Южная Австралия, Австралия
4
Photonics Engineering Group (GIF), Департамент TEISA, Университет Кантабрии, E-39005 Сантандер, Испания
5
CIBER-bbn, Instituto de Salud Carlos III, E-28029 Мадрид, Испания
6
IDIVAL, Instituto de Investigación Marques Valdecilla, E-39011 Сантандер, Кантабрия, Испания
7
Пекинский научно-исследовательский центр прикладных лазерных технологий; Институт лазерной инженерии, Пекинский технологический университет, Пекин 100124, Китайская Народная Республика
8
Кафедра электротехники, Гонконгский политехнический университет, Коулун, Гонконг
9
Лаборатория лазерной инженерии и приложений, факультет электротехники и вычислительной техники, Сеульский национальный университет, Сеул 08826, Республика Корея
10
Max-Planck-Institut für Quantenoptik, Hans-Kopfermann-Str.1. D-85748 Гархинг, Германия
11
Государственная ключевая лаборатория современных оптических приборов, Колледж оптических наук и инженерии, Чжэцзянский университет, Ханчжоу 310027, Китайская Народная Республика
12
Центр фотоники, Бостонский университет, 8 Saint Mary’s Street, Бостон, Массачусетс 02215, Соединенные Штаты Америки
13
Химический факультет, Бостонский университет, 590 Commonwealth Avenue, Бостон, Массачусетс 02215, Соединенные Штаты Америки
14
RDRL-SEE-O, Исследовательская лаборатория армии США, 2800 Powder Mill Road, Adelphi, Maryland 20783, Соединенные Штаты Америки
15
MCH Engineering LLC, Аламеда, Калифорния 94501, Соединенные Штаты Америки
16
Лаборатория спектральной диагностики, факультет химии и химической биологии, Северо-Восточный университет, Бостон, Массачусетс 02115, Соединенные Штаты Америки
17
Cireca Theranostics, LLC, Кембридж, Массачусетс 02139, Соединенные Штаты Америки
18
Институт живых систем, факультет физики и астрономии, Эксетерский университет, Эксетер, EX4 4QD, Соединенное Королевство
19
Институт Роуленда при Гарвардском университете, Кембридж, Массачусетс 02142, Соединенные Штаты Америки
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере. - Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с вашим системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Перейти к основному содержанию
Поиск
Поиск
- Где угодно
Быстрый поиск где угодно
Поиск JCSMSearch
Расширенный поиск
Авторизоваться
Пропустить основную навигацию Закрыть меню ящика Открыть меню ящика Меню
- На главную
- Проблемы
- Текущий
- Архив
- Принятые статьи
- RSS-канал
- Коллекции
Стандарты практики
- EM : Резиденты / стипендиаты
- COVID-19
- 15-летний юбилей
- Преимущества для авторов
- Правила подачи заявок
- О JCSM
- Совет директоров
- Перепечатки и разрешения
- Рецензенты
- Рекламодатели
- Политика журнала
- Подписка
Проблемы
Проблемы
- Текущий
- Архив
- Принятые статьи
- Новостная лента
Коллекции
Коллекции
- Стандарты практики
- REM: резиденты / стипендиаты
- COVID-19
- 15-летний юбилей
Отправить
Отправить
- Преимущества для авторов
- Правила подачи документов
Информация
Информация
- О JCSM
- Редакция журнала
- Заработайте CME
- Перепечатки и разрешения
- Рецензенты
- Рекламодатели
- Правила журнала
- Подписаться
Подпишитесь на нас в
Твиттер
RSS
© 2021 Американская академия медицины сна
Условия эксплуатации
Члены Американской академии медицины сна
Войти через аккаунт AASM
Американский альянс за здоровый сон участников
Войдите через учетную запись AAHS
Путь анемии Фанкони и восстановление ICL: значение для лечения рака
Abstract
Анемия Фанкони (FA) — это наследственное заболевание, вызываемое мутациями по крайней мере в 13 генах и характеризующееся нестабильностью генома.Помимо демонстрации поразительно гетерогенных клинических фенотипов, пациенты с FA чрезвычайно чувствительны к лечению с помощью сшивающих агентов, которые создают межнитевые сшивки (ICL). В отличие от бактерий и дрожжей, у которых ICL репарируются посредством зависимых от репликации и независимых механизмов, считается, что ICL репарируются в первую очередь во время репликации ДНК у позвоночных (Moldovan and D’Andrea, 2009). Однако недавние данные показывают, что независимая от репликации репарация ICL также действует у позвоночных.Хотя точная роль пути FA в репарации ICL остается неуловимой, все больше данных свидетельствует о том, что белки FA функционируют на разных этапах восприятия, распознавания и обработки ICL, а также в передаче сигналов от этих очень токсичных повреждений, которые могут быть вызваны широкий выбор химиотерапевтических препаратов против рака. Здесь мы обсуждаем некоторые из недавних открытий, которые пролили свет на роль пути FA в восстановлении ICL, с особым акцентом на значение этих открытий для терапии рака, поскольку нарушение генов FA было связано с предрасположенностью к раку.
Ключевые термины: репарация ДНК, репликация ДНК, перекрестная связь, контрольная точка, анемия Фанкони
Введение
Анемия Фанкони (FA) — редкое наследственное заболевание, связанное с нестабильностью генома, которое привлекло внимание широкой аудитории со стороны фундаментальных ученых изучение механизмов восстановления ДНК для онкологов, занимающихся лечением различных спорадических форм рака. Пациенты с FA характеризуются аномалиями развития, прогрессирующей недостаточностью костного мозга и предрасположенностью к раку, особенно лейкозам и карциномам.Несмотря на разнообразные клинические фенотипы заболевания, отличительной чертой клеток, полученных от пациентов с FA, является тяжелая клеточная гиперчувствительность к сшивающим агентам, таким как митомицин C (MMC), цисплатин (CDDP) и диэпоксибутан (DEB) (см. Обзор в (Moldovan and D. Andrea, 2009; Niedernhofer et al., 2005; Patel, Joenje, 2007; Thompson, Hinz, 2009; Wang, 2007)). Эта характеристика привела к большому количеству исследований, касающихся участия пути FA в репарации поперечных сшивок и поддержании стабильности генома.Кроме того, путь FA также был связан с восприимчивостью к раку, в первую очередь с опухолями груди, а также с чувствительностью и устойчивостью к химиотерапевтическим агентам. Следовательно, лучшее понимание механизмов и роли пути FA не только поможет пациентам, страдающим этим редким заболеванием, но также окажет влияние на онкологических больных в общей популяции.
Путь FA
Анемия Фанкони возникает в результате мутаций в одном из 13 генов FA: Fanca , -b , -c , -d1 , -d2 , -e , — f , -g , -i , -j , -l , -m и -n ().FA-ассоциированные мутации являются аутосомно-рецессивными, за исключением Fancb , который является X-сцепленным. Белки, кодируемые этими генами, составляют путь ЖК. Путь FA можно разделить на три группы: основной комплекс FA, комплекс FANCD2 / FANCI (ID) и белки FA, действующие ниже по течению. Для более подробных обзоров пути FA см. (Kennedy and D’Andrea, 2005; Levitus et al., 2006; Mathew, 2006; Taniguchi and D’Andrea, 2006). Ядровый комплекс FA состоит из FANC-A, -B, -C, -E, -F, -G, -L и -M, а также дополнительных белков FAAP24 и FAAP100.Нижестоящие белки FA состоят из FANCD1, FANCJ и FANCN, которые также известны как BRCA2, BRIP1 / BACh2 и PALB2 соответственно. FANCD1 / BRCA2 является основным геном предрасположенности к раку груди и яичников и играет важную роль в гомологически направленном восстановлении (HDR) (Godthelp et al., 2006a; Godthelp et al., 2006b). FANCJ взаимодействует напрямую с BRCA1 и является членом семейств DEAH и XPD геликаз (Cantor et al., 2001; White, 2009). FANCN взаимодействует с FANCD1 / BRCA2 и требуется для его функций рекомбинации и контрольных точек (Xia et al., 2006a; Xia et al., 2006b). Эта поразительная связь между путем FA и восприимчивостью к раку груди, по-видимому, ограничивается этим субнабором генов FA (Garcia et al., 2009). Путь FA является уникальным примером, в котором мутации из большой эпистатической группы генов связаны с одним и тем же заболеванием. Недавно гомозиготная мутация RAD51C, паралога RAD51, была идентифицирована в семье с множественными врожденными аномалиями, характерными для FA (Vaz et al., 2010). Основываясь на эффекте потери функции RAD51C на образование фокусов RAD51, RAD51C, по-видимому, действует ниже комплекса ID.
Таблица 1
Белки Фанкони, функции и связи с раком.
| Белок FA | Субкомплекс | Роль | Связь с раком | |||
|---|---|---|---|---|---|---|
| FANCA | Core Complex | D2 / I80, активированный с помощью инактивации 904, фосфорилированный 904 при инактивации AT, фосфорилированный 904 | ||||
| FANCB | Core Complex | Убиквитилирование D2 / I | ||||
| FANCC | Core Complex | D2 / I Убиквитилирование | D2 / I | Инактивировано с помощью промотора 9047 AMLANC 904 904 Комплекс | Убиквитилирование D2 / I, фосфорилированное при повреждении (CHK1) | |
| FANCF | Базовый комплекс | Убиквитилирование D2 / I | Инактивировано (метилирование) при раке яичников и легких | |||
| FANCG | Core Complex | Убиквитилирование D2 / I, фосф. орилированный в M | ||||
| FANCL | Core Complex | E3-лигаза для комплекса D2 / I | Измененная экспрессия в вариантах сплайсинга | |||
| FANCM | Core Complex Локализация в ДНК Pocphase | |||||
| FANCD2 | Убиквитилированный комплекс | Убиквитилированный и фосфорилированный при повреждении (ATM, ATR, CHK1) | ||||
| ATRI | Убиквитилированный | Убиквитилированный комплекс | ||||
| FANCD1 / BRCA2 | Нижестоящие очаги | HR-компонент | Восприимчивость к раку молочной железы, яичников, предстательной железы, поджелудочной железы, чувствительность к цисплатину | |||
| FANCJ / BACh2 | FANCJ / BACh2 | FANCJ / BACh 2 | ||||
| FANCN / PALB2 | Downstre am foci | Связывание FANCD1 | Восприимчивость к раку молочной железы.Инактивировано метилированием промотора |
Модификации белков FA: убиквитилирование
Центральным элементом функции и регуляции пути FA является убиквитилирование комплекса FANCD2 / FANCI. Основной комплекс FA моноубиквитилирует FANCD2 и FANCI и активирует путь FA (Ciccia et al., 2007; Dorsman et al., 2007; Garcia-Higuera et al., 2001; Ling et al., 2007; Meetei et al., 2003). ; Sims et al., 2007). В то время как FANCL является каталитической субъединицей (Meetei et al., 2003), все десять членов основного комплекса FA необходимы для его активности лигазы убиквитина E3, а UBE2T служит лигазой убиквитина E2 (Alpi et al., 2007; Alpi et al., 2008; Machida et al., 2006) . FANCM, однако, можно отличить от других компонентов основного комплекса, поскольку FANCD2 все еще частично убиквитилирован, а основной комплекс остается интактным после инактивации FANCM (Kim et al., 2008). Это привело к предположению, что FANCM-FAAP24 представляет собой субъединицу, нацеленную на ядерный комплекс ДНК (Ali et al., 2009; Ciccia et al., 2007). Кроме того, FANCM и FAAP24, как было показано, взаимодействуют с HCLK2 независимо от комплекса ядра FA и функционируют в передаче сигналов контрольных точек, опосредованной ATR (Collis et al., 2008). Другой вспомогательный белок, FAAP100, как было показано, взаимодействует в стабильном комплексе с FANCL и FANCB и важен для моноубиквитилирования FANCD2 и стабильности ядерного комплекса FA (Ling et al., 2007). Моноубиквитилирование FANCD2 и FANCI имеет решающее значение для активации пути FA, поскольку оно требуется для образования и локализации комплекса FANCI / D2 в ядерных фокусах (Dorsman et al., 2007; Sims et al., 2007; Smogorzewska et al., 2007; Ван, 2007). Функциональное группирование белков FA было выведено из их роли в убиквитилировании FANCD2 / FANCI (). Все компоненты основного комплекса требуются для модификации идентификатора и, следовательно, находятся в восходящем направлении. Инактивация нижележащих белков FA не влияет на убиквитилирование ID. Комплекс ID образован моноубиквитилированными FANCD2 и FANCI, чьи моноубиквитилирования взаимозависимы (Smogorzewska et al., 2007). Комплекс ID образуется и локализуется в хроматине во время S-фазы (Taniguchi et al., 2002a) и в ответ на агенты, повреждающие ДНК, включая MMC, ионизирующее излучение, ультрафиолетовый свет и гидроксимочевину (Garcia-Higuera et al., 2001). На ДНК комплекс ID образует фокусы с белками репарации ДНК BRCA1 и RAD51 и нижележащими белками FA (Garcia-Higuera et al., 2001; Taniguchi et al., 2002a; Wang et al., 2004). В то время как убиквитилирование FANCD2 по K561 важно для активации пути FA, убиквитилирование FANCI по K523 — нет (Ishiai et al., 2008).
Путь FA: субкомплексы, белки и посттрансляционные модификации
Гипотетическая локализация различных субкомплексов FA в репликационной вилке, сталкивающейся с межцепочечной перекрестной связью (красная линия).Основной комплекс вместе с E2-лигазой UBE2, моноубиквитилатом FANCI и FANCD2 (комплекс ID), запускает его ассоциацию с поврежденным хроматином. Некоторые белки FA фосфорилируются в ответ на повреждение ДНК киназами контрольной точки ATM, ATR и CHK1. Кроме того, FANCM и FANCG фосфорилируются в митозе (подробности см. В тексте). Примечание: цветная версия рисунка доступна в Интернете.
Комплекс USP1 / UAF1 деубиквитилирует FANCD2 (Cohn et al., 2007; Nijman et al., 2005) и FANCI (Longerich et al., 2009). Деубиквитилирование FANCD2 необходимо для восстановления ICL (Oestergaard et al., 2007), а потеря USP1 связана с чувствительностью ICL (Kim et al., 2009). Примечательно, что инактивация USP1 у мышей воспроизводит несколько фенотипов FA, напоминающих нокаут генов FA (Kim et al., 2009). Ингибиторы протеасом повышают чувствительность клеток к повреждению ДНК, и потенциальной мишенью для этой сенсибилизации является активация пути FA (Jacquemont and Taniguchi, 2007). Однако конкретный белок-мишень протеасомы не идентифицирован.
Модификации белков ЖК: фосфорилирование
Некоторые белки ЖК фосфорилируются (;). Эти модификации регулируют активность пути FA после повреждения ДНК и во время клеточного цикла. В то время как фосфорилирование белков FA обычно происходит в ответ как на повреждение ДНК, так и на стресс репликации, FANCA специфически фосфорилируется с помощью ATR (связанного с ATM и Rad3) после повреждения ДНК, но не во время репликации ДНК (Collins et al., 2009). FANCM фосфорилируется в ответ на репликацию ДНК и повреждение ДНК, и это фосфорилирование регулируется ATM PIKKs (мутированная атаксия телеангиэктазии) и ATR (Kim et al., 2008; Собек и др., 2009). Последствия зависимого от повреждения ДНК фосфорилирования FANCM неизвестны. FANCD2 фосфорилируется по множеству сайтов после повреждения ДНК. Фосфорилирование с помощью ATR является критическим для передачи сигналов контрольных точек во время S-фазы (Andreassen et al., 2004) и для моноубиквитилирования FANCD2 (Ho et al., 2006). FANCD2 также фосфорилируется ATM; однако эта модификация не зависит от моноубиквитилирования FANCD2 (Taniguchi et al., 2002b). FANCD2 и FANCE фосфорилируются с помощью CHK1, и эти модификации играют роль в устойчивости к сшивающим агентам ДНК (Wang et al., 2007; Zhi et al., 2009). Наконец, FANCG и FANCM гиперфосфорилируются во время митоза (Kee et al., 2009; Mi et al., 2004). Хотя роль этих зависимых от клеточного цикла фосфорилирования неизвестна, они могут отражать потенциальную роль пути FA в митозе. Примечательно, что полногеномный скрининг siRNA генов, которые регулируют прогрессирование митоза, показал, что подавление FANCC и FANCE приводит к задержке митоза (Neumann et al., 2010). Кроме того, FANCD2 локализуется в ультратонких мостах хромосом во время анафазы, где он локализуется совместно с BLM (Neumann et al., 2010), подтверждая возможную роль FANCD2 в предотвращении анеуплоидии (Naim and Rosselli, 2009).
Сшивающие агенты и межцепочечные сшивки ДНК
ICL являются особенно токсичными повреждениями ДНК, потому что они затрагивают обе цепи ДНК, блокируя важные процессы, которые требуют транслокации вдоль ДНК; а именно репликация и транскрипция ДНК. В дополнение к этому физическому ограничению ДНК, ICL требуют восстановления повреждений обеих цепей ДНК. В сравнительных исследованиях, оценивающих генотоксичность in vitro, и in vivo, больших наборов соединений, сшивающие агенты были отнесены к числу наиболее токсичных (Lohman, 1999).Сшивающие агенты ДНК вызывают грубые хромосомные аберрации, включая потерю хромосом, делеции и разрывы, а способность вызывать хромосомные аберрации коррелирует с их цитотоксичностью (Vogel et al., 1998; Vogel et al., 1996). Следовательно, сшивающие агенты — это в первую очередь кластогены, а не мутагены.
ICL могут быть обнаружены различными механизмами в зависимости от типа транзакции ДНК, происходящей в сшитой молекуле ДНК. В активно делящихся клетках репликативный аппарат будет сталкиваться с ICL во время S-фазы, и ICL будет действовать как физический барьер для развития реплисом.Считается, что это распространенный механизм обнаружения повреждений ICL. В этом случае восстановление ICL включает в себя столкновение вилки репликации в качестве триггера для инициации восстановления. При обнаружении ICL во время репликации ДНК активация контрольной точки, многоступенчатая репарация и стабилизация реплисомы должны быть тесно скоординированы. Восстановление ICL у позвоночных включает образование хрупких промежуточных звеньев ДНК, включая DSB, разрывы и разрывы оцДНК. Если эти структуры ДНК оставить нереставрированными или частично восстановленными, они могут привести к коллапсу репликационной вилки и событиям аберрантной рекомбинации, таким как слияние хромосом, которое приводит к радиальным хромосомам, наблюдаемым у пациентов с FA.
Предполагается, что этого зависимого от репликации режима репарации должно быть достаточно, чтобы справиться с ICL и своевременно восстановить ее. Однако в непролиферирующих клетках, таких как постмитотические дифференцированные клетки или покоящиеся стволовые клетки, эндогенно сгенерированные ICL необходимо восстанавливать в отсутствие репликации ДНК. В этой ситуации, если ICL расположен в активно транскрибируемом гене, подобный процесс восприятия может иметь место после остановки РНК-полимеразы, и блок транскрипции может быть инициирующим событием.Однако гораздо меньше известно о последствиях блокирования транскрипции ICL. Наконец, считается, что искажающие ICL () могут быть распознаны в отсутствие столкновения с репликационным или транскрипционным механизмом, движущимся вдоль ДНК.
Таблица 2
Свойства различных классов сшивающих агентов. Нуклеотиды, выделенные жирным шрифтом, показывают сшитые нуклеотиды в ICL.
| Азотные иприты | Соединения платины | Нитрозомочевины | Митомицин C | Псоралены | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C | BCMC | C | Псоларен, 8-Метоксипсорален | |||||||
| Сшитые последовательности (ICL) | 5′- G NC-3 ‘ 3′-CN G -5′ | 5′- G C-3 ‘ 3′-C G -5 ‘ | 5′- G -3′ 3′- C -3 ‘ | 5′- C G-3′ 3′-G C -5 ‘ | 5′- T A-3′ 3′-A T -5 ‘ | |||||
| Искажение | Переменное | Основное + изгиб | Неизвестно | Скромное Основное | ||||||
| Доля ICL | 1–5% | 2–5% | 3–8% | 5–13% | До 40% | |||||
| Другие основные поражения | Моноаддукты | Внутрирядные сшивки | Сшивки внутри цепей | Моно-аддукты | Моноаддукты, АФК | Моноаддукты | ||||
| Химиотерапевтические применения | Лимфоидные опухоли | Яичники, яички, легкие | Глиобластома | Гастроинтестинация, молочная железа | Желудочно-кишечный тракт, грудная клетка, мочевой пузырь | (Dy and Adjei, 2008; Rink and Hopkins, 1995) | (Coste et al., 1999; Хуанг и др., 1995; Kartalou and Essigmann, 2001) | (Colvin et al., 1976; Wiencke and Wiemels, 1995) | (Pagano, 2002; Rajski and Williams, 1998; Warren et al., 1998) | (Averbeck et al., 1988; Honig et al., 1994; Spielmann et al., 1995; Stern, 2007) |
Сшивающие агенты и их актуальность
ICL генерируются бифункциональными агентами, которые относятся к разным классам. Азотные горчицы (бис (2-хлорэтил) метиламин: HN2, хлорамбуцил, мелфалан) используются более 65 лет и до сих пор широко используются для лечения рака (Gilman and Philips, 1946), в частности, для лимфоидных опухолей (множественная миелома, хронический лимфолейкоз (ХЛЛ)).Они реагируют с N7 гуанина с образованием в основном моноаддуктов к ДНК. Однако 1–5% поражений представляют собой ICL, которые ответственны за цитотоксичность этих соединений (Rink and Hopkins, 1995).
Цисплатин (цисплатиндиамминхлорид II, CDDP) и ди- и трехъядерные аналоги цис -DDP были впервые идентифицированы как ингибиторы бактериального деления (Rosenberg et al., 1965) и используются для лечения широкого спектра солидных опухолей с Наибольший эффект наблюдается при лечении рака яичек и яичников.Как и в случае с азотным ипритом, ICL представляют собой небольшую часть повреждений, вызванных препаратами платины (1–5%), которые в основном представляют собой поперечные связи внутри нитей (Brabec, Ленг, 1993; Jones et al., 1991; Kartalou and Essigmann, 2001). Эти соединения также реагируют с N7 гуанина с образованием ICL. Хотя считается, что ICL являются основной причиной цитотоксичности (Roberts and Friedlos, 1987), стоит отметить, что внутрицепочечные перекрестные связи — это повреждения, которые запускают контрольную точку и активируют пути репарации ДНК, перекрывающиеся с репарацией ICL.Примечательно, что сообщалось о различиях в цитотоксичности и эффективности между соединениями платины, образующими отчетливые внутрицепочечные сшивки. Напр., Транс-DDP, менее токсичный аналог цис-DDP, не может образовывать внутрицепочечные сшивки 5′-GG-3 ‘(Aris and Farrell, 2009).
Митомицин С (MMC) — это антибиотик из группы Streptomyces , который широко используется в клеточных исследованиях ICL (Tomasz, 1995). MMC необходимо метаболизировать до активных промежуточных продуктов, которые запускают 5–14% ICL, среди других аддуктов (Tomasz, 1995).MMC также генерирует активные формы кислорода (ROS), которые способствуют его токсичности (Pagano, 2002). MMC используется для лечения рака желудочно-кишечного тракта, груди, легких и мочевого пузыря. Другой класс антибиотиков из Streptomyces — это пирроло [2,1-c] [1,4] бензодиазепины (PBD), которые очень эффективно образуют ICL (Clingen et al., 2005; Gregson et al., 2001).
Нитрозомочевины (BCNU, кармустин) представляют собой химиотерапевтические агенты, которые метаболизируются до бифункциональных молекул, которые, в свою очередь, генерируют менее 8% ICL среди других аддуктов (Wiencke and Wiemels, 1995).Однако токсичность ограничивает клиническое использование нитрозомочевины.
Наконец, псорален и его производные образуют межцепочечные сшивки с ДНК при активации УФ-излучением длинноволновым УФ-светом. Псоралены используются для лечения псориаза, витилиго и кожной Т-клеточной лимфомы. Они образуют самую высокую долю ICL (до 40%) среди сшивающих агентов (Averbeck et al., 1988; Dronkert and Kanaar, 2001; Honig et al., 1994; Warren et al., 1998).
Пути репарации ICL в первую очередь предполагаются благодаря использованию этих сшивающих агентов.Однако различные синтетические ICL также использовались для исследований in vitro и исследований бесклеточных экстрактов (см. Ниже). Разработка систем, использующих шаблоны ДНК, несущие одиночные, определенные повреждения ICL, дает существенное понимание механизмов обнаружения, передачи сигналов и восстановления ICL. Эта стратегия является значительным прогрессом по сравнению с использованием сшивающих агентов, которые генерируют только меньшую часть из межцепочечных перекрестных связей и в основном генерируют внутрицепочечных перекрестных связей и другие формы повреждения ДНК, которые способны активировать передачу сигналов контрольной точки и активировать механизмы восстановления ().
Было показано, что ICL могут возникать в клетках эндогенно (Niedernhofer et al., 2003). Предлагаемые источники эндогенных ICL включают альдегиды, такие как малоновый диальдегид (Kozekov et al., 2003; Stone et al., 2008). Эти соединения, которые действуют по существу как бифункциональные алкилирующие агенты, являются продуктами перекисного окисления липидов. Другой источник ICL — отслеживание действия связанных с псораленом фурокумаринов, которые могут быть обнаружены в растениях (Kleiner et al., 2003). Степень образования эндогенных ICL в клеточной ДНК и их возможные биологические эффекты остаются неясными, однако был сделан вывод, что до 10 ICL могут генерироваться и восстанавливаться на клетку каждый день (Lindahl and Barnes, 2000).Накопление повреждений ДНК ICL со временем может способствовать старению клеток (Grillari et al., 2007).
Наиболее убедительные доказательства, подтверждающие идею о том, что ICL представляют собой токсические поражения среди аддуктов ДНК, генерируемых сшивающими агентами, получены из сравнительных исследований между бифункциональными соединениями, которые способны образовывать ICL, и их химически родственными монофункциональными аналогами, которые ограничены образованием моноаддуктов. . В то время как монофункциональные алкилирующие агенты в основном вызывают точечные мутации, которые легко объяснить типом образующегося продукта алкилирования, бифункциональные сшивающие агенты вызывают серьезные хромосомные изменения.Например, исследования, сравнивающие хлорамбуцил с его монофункциональным аналогом, показывают, что хотя хлорамбуцил, азотистый иприт, вызывающий образование ICL, лечение приводило к делециям, его монофункциональный аналог вызывал точечные мутации (Yaghi et al., 1998). Более того, агенты, генерирующие ICL, более эффективны в индукции хромосомных потерь и обменов сестринских хроматид, чем их монофункциональные аналоги (Bodell et al., 1985; Vogel et al., 1996).
обобщает характеристики некоторых сшивающих агентов, используемых в терапии рака и / или для экспериментальных исследований ICL.Примечательно, что разные источники сшивания дают разные диапазоны аберрантных структур ДНК, из которых ICL всегда составляют менее многочисленную фракцию. Более того, ICL обладают уникальными структурными характеристиками. В частности, физико-химическая природа ICL влияет на топологию окружающей спирали ДНК, которая, в свою очередь, как полагают, влияет на способ распознавания и восстановления ICL (Noll et al., 2006; Rajski and Williams, 1998 ; Смитон и др., 2008; Смитон и др., 2009).
Модельные системы для изучения восстановления ICL
Некоторые характеристики различных стратегий восстановления ICL описаны ниже.Более полные обзоры см. В (Dronkert and Kanaar, 2001; McVey, 2010; Niedernhofer et al., 2005; Patel and Joenje, 2007). Восстановление ICL было впервые исследовано на E. coli (Cole, 1973). Там восстановление ICL начинается с разрезов по обе стороны от ICL посредством Uvr (A) 2BC (Sladek et al., 1989; Van Houten et al., 1988). Если доступна гомологичная рекомбинация, резекция в одном из разорванных положений вызывает загрузку RecA и инвазию неповрежденной матрицы с последующим синтезом ДНК в области, содержащей поперечную сшивку.Затем Uvr (A) 2BC удаляет незакрепленную сшивку, и образовавшийся промежуток заполняется. Когда HR недоступен, после разрезов Uvr (A) 2BC происходит подверженный ошибкам трансфузионный синтез (через Pol II). Для восстановления ICL у дрожжей используются сходные стратегии, но, кроме того, он строго регулируется клеточным циклом (McHugh et al., 2001). Следует отметить, что модели репарации ICL в S. cerevisiae в основном основаны на генетических данных, и последовательность событий остается гипотетической. Во время S-фазы конфликт репликационной вилки с ICL приводит к генерации DSB (McHugh et al., 2000). Этот DSB может возникать как следствие нуклеолитического расщепления после остановки вилки из-за ICL или может быть результатом встречи вилки с однонитевым разрезом, образованным ранее существовавшим разрезом в непосредственной близости от ICL. Затем ICL отсоединяется и после синтеза трансфузии удаляется путем эксцизионной репарации нуклеотидов (NER). Кроме того, путь HR необходим для репарации промежуточного соединения DSB, что убедительно подтверждается чувствительностью мутантов HR к сшивающим агентам (Henriques and Moustacchi, 1981).Напротив, в клетках G1 отсутствуют движущиеся репликационные вилки для восприятия ICL и отсутствуют реплицированные гомологичные последовательности для использования на этапе HR при репарации. В G1 распознавание ICL зависит от аппарата NER (Lehoczky et al., 2007) и сопровождается синтезом трансфузии и вторым набором разрезов, высвобождающих перекрестно-связанный нуклеотид. Возникновение репарации ICL в G1 у дрожжей подтверждается мутагенной природой репарации и чувствительностью дрожжевых мутаций в подверженной ошибкам пострепликационной репарации к сшивающим агентам (Henriques and Mouracchi, 1981; McHugh et al., 2000). Примечательно, что дрожжи используют разные наборы путей восстановления для восстановления повреждений, вызванных разными сшивающими агентами (Beljanski et al., 2004). Это говорит о том, что структура сшивки влияет, по крайней мере частично, на способ ремонта.
Восстановление ICL у высших эукариот менее изучено. Однако считается, что между дрожжами и высшими эукариотами существует некоторое фундаментальное сходство. Восстановление ICL сопровождается образованием DSB, для восстановления которого требуется HR. Хотя HR- и репликационно-зависимая репарация ICL считается основным способом репарации, описаны также независимые от репликации, HR-независимые и подверженные ошибкам пути репарации ICL (Wang et al., 2001; Zheng et al., 2003). В частности, было показано, что поперечные связи псоралена, индуцированные УФ-лазерным облучением, восстанавливаются в клетках G1 с помощью XPC-зависимого механизма репарации (Muniandy et al., 2009). Кроме того, с помощью количественной ОТ-ПЦР для репарированных повреждений было показано, что плазмида, несущая единственную ICL, репарируется в G1-арестованных клетках млекопитающих ((Ben-Yehoyada et al., 2009) и см. Ниже).
Принципиальным отличием дрожжей от других организмов является потребность в белках ЖК.Хеликазы FANCJ и FANCM появились первыми в эволюции и имеют гомологов у бактерий и архей (Meetei et al., 2005; White, 2009). У беспозвоночных есть несколько генов FA с комплексом FANCD2 / I и нижестоящими генами FA: FANCD1 и FANCJ у C. elegans и FANCD1 у Drosophila . В то время как Drosophila имеет гомолог FANCL, C. elegans , по-видимому, лишен белков ядра комплекса FA, что ставит под вопрос идентичность убиквитинлигазы, ответственной за убиквитилирование FANCD2, которое наблюдается у C.elegans (McVey, 2010).
Большинство исследований репарации FA и ICL было выполнено на клетках человека и мыши, а также на моделях мышей и лимфобластных клетках курицы (DT40). Кроме того, бесклеточные экстракты яиц Xenopus недавно стали мощной системой для изучения восстановления ICL и передачи сигналов от ICL.
белков FA в ответе на повреждение ICL
белков FA в качестве сенсоров повреждения ДНК
Путь FA участвует в ответе на повреждение ДНК, включая, помимо прочего, повреждение ICL.Некоторые белки FA модифицируются после повреждения ДНК, включая моноубиквитилирование FANCD2 и FANCI (). Было высказано предположение, что помимо их роли ниже киназ контрольных точек, белки FA могут также играть роль на ранних этапах реакции на повреждение ДНК, то есть в качестве сенсоров повреждения ДНК.
Было показано, что путь FA является ранним ответчиком на повреждение ICL. Клетки FA обнаруживают дефект на начальном этапе разреза ICL, обычно связанный с активностью XPF / ERCC1 (Fujiwara, 1982; Kumaresan and Lambert, 2000; Kumaresan et al., 2007), хотя XPF может иметь дополнительные функции (см. Обзор в (Bergstralh and Sekelsky, 2008; Hlavin et al., 2010)). Это вовлекает путь FA в восприятие ICL и / или рекрутирование XPF / ERCC1. Эта идея была недавно подтверждена экспериментами с бесклеточными экстрактами Xenopus, показавшими, что истощение FANCD2 приводит к дефекту разреза и последующей задержке синтеза трансфузии (Knipscheer et al., 2009).
Эксперименты с использованием небольших плазмидных матриц, несущих одну ICL, значительно укрепили идею о том, что белки FA могут играть прямую роль в обнаружении ICL.Шен и др. использовали субстрат, несущий сайт начала репликации EBV и индуцированную псораленом ICL, и обнаружили, что белки FA обогащены в сайте ICL (Shen et al., 2009). Важно, что в этих экспериментальных условиях компоненты ядра комплекса FA и FANCD2 / FANCI рекрутируются в ICL независимым от репликации способом, вовлекая путь FA на ранней стадии восприятия ICL. Напротив, рекрутирование нижележащих белков FA FANCD1, FANCJ и FANCN требует репликации, но не базового комплекса FA, что позволяет предположить, что ядро FA и белки FANCD2 / FANCI рекрутируются непосредственно в ICL, а связанные с BRCA белки FA рекрутируются в остановка репликационной вилки в ICL (Shen et al., 2009). Дифференциальное привлечение белков FA может отражать участие пути FA в различных путях репарации ICL и согласуется с сенсорной ролью белков FA в независимой от репликации репарации ICL и ролью нижележащих белков FA в HDR, которая требует место в репликационно-зависимой репарации ICL. В соответствии с этим, используя бесклеточные экстракты Xenopus, было показано, что белки FA рекрутируются в плазмиду, несущую единственную искажающую ICL, независимо от того, имеет место репликация ДНК или нет (Ben-Yehoyada et al., 2009). Также было продемонстрировано, что рекрутирование белков FA не зависит от RPA- и ATR-зависимых контрольных точек (Ben-Yehoyada et al., 2009). Эти данные указывают на путь FA на ранней стадии восприятия ICL, выше передачи сигналов контрольной точки. Это может объяснить, почему клетки FA настолько чувствительны к ICL, поскольку они неспособны правильно распознавать ICL и, следовательно, не могут активировать последующие необходимые шаги для удаления ICL.
Белки FA и восстановление ICL
Повышенная чувствительность к сшивающим агентам, присущая всем пациентам с FA, указывает на то, что путь FA играет важную роль в восприятии, передаче сигналов или восстановлении повреждений, вызванных этими агентами.Однако точная роль белков FA в ответе на ICL все еще остается в значительной степени неуловимой. Путь FA может участвовать как в зависимых от репликации, так и в независимых от репликации путях репарации ICL, описанных выше. Когда клетки FA подвергаются воздействию сшивающих агентов, они накапливают хромосомные разрывы и радиальные хромосомы (Auerbach, 1993), что указывает на дефект клеточного ответа на ICL.
После распознавания ICL и остановки клеточного цикла передачи сигналов, позволяющей его репарацию, путь FA может затем функционировать, чтобы координировать репарацию ICL.Нарушение основного комплекса FA и комплекса ID, как было показано, снижает эффективность восстановления ICL (Ben-Yehoyada et al., 2009; Shen et al., 2009). Считается, что механизмы репарации ICL включают HDR, синтез трансформации (TLS) и часть механизма NER. В то время как TLS и нуклеаза (ы) необходимы как для зависимой от репликации, так и для независимой от репликации репарации, считается, что зависимая от репликации репарация необходима только при репликационно-зависимой репарации, когда доступны последовательности, гомологичные повреждению.Однако также возможно, что независимая от репликации, HDR-зависимая репарация ICL может действовать в фазе G2 клеточного цикла. Примечательно, что путь FA был связан с белками, участвующими в HDR, TLS и NER. Точная роль белков FA в HDR неясна. В то время как клетки FANCD1 обнаруживают заметный дефект HDR (Moynahan et al., 2001; Nakanishi et al., 2005; Stark et al., 2004), другие клетки FA демонстрируют более умеренный дефект HDR (Hirano et al., 2005; Nakanishi et al., 2004). al., 2005; Niedzwiedz et al., 2004; Yamamoto et al., 2005; Ямамото и др., 2003; Ян и др., 2005). Однако связь между путем FA и путем BRCA хорошо установлена, поскольку FANCD1 идентичен BRCA2 и несколько белков пути FA взаимодействуют непосредственно с BRCA1 или BRCA2. Следовательно, возможно, что хотя путь FA не играет главной роли во всех HDR, он играет специфическую роль в привлечении репарационных белков и координации репарации HDR в контексте повреждения ICL. Действительно, in vitro, исследования показывают, что FANCD1 / BRCA2 играют важную роль в репарации ICL (Cipak et al., 2006). FANCD2 также ассоциирует с комплексом Mre11-Rad50-Nbs1 (MRN), который важен для резекции ДНК в DSBs, первой стадии всех гомологически зависимых процессов репарации (Roques et al., 2009). MRN необходим для образования FANCD2 фокусов в сайтах повреждения: пробелах ssDNA и DSBs (Roques et al., 2009).
Было показано, что в ответ на сшивающие агенты FANCD2 колокализуется с компонентом NER XPF (Sridharan et al., 2003), а XPF влияет на стабильность убиквитилированного FANCD2 (Bhagwat et al., 2009). FANCD2 также, как было показано, колокализуется с TLS полимеразой Rev1 после ареста репликации (Niedzwiedz et al., 2004), а компоненты ядра комплекса FA, как было показано, необходимы для эффективного образования фокусов Rev1 (Mirchandani et al., 2008). Играя специфическую роль ICL в HDR и роль выше TLS и NER, путь FA помогает координировать и регулировать эти механизмы репарации для эффективного и правильного устранения повреждений ICL. В соответствии с этими идеями было показано, что инактивация FANCD2 влияет как на нуклеолитический разрез, так и на синтез трансфузии (Knipscheer et al., 2009).
Недавние эксперименты изучали роль пути FA в репарации ICL с использованием ДНК-субстратов, содержащих одну сайт-специфичную ICL в бесклеточных экстрактах Xenopus (Ben-Yehoyada et al., 2009; Raschle et al., 2008) . Кроме того, поведение ICL-содержащих ДНК-матриц отслеживали в клетках млекопитающих после трансфекции (Ben-Yehoyada et al., 2009; Shen et al., 2009).
В одном из исследований Xenopus сообщается о системе, в которой восстановление ICL зависит исключительно от репликации (Knipscheer et al., 2009; Raschle et al., 2008). Это исследование подробно описывает начальные шаги в процессинге ICL в репликационно-зависимой системе с использованием прежде всего азотистого иприта-подобного аддукта ICL, который минимально искажает спираль ДНК (Raschle et al., 2008). Две репликационные вилки сходятся на ICL, при этом ведущая цепь задерживает первоначально 20-24 нуклеотида от ICL с последующим продвижением одной ведущей цепи в пределах 1 нуклеотида от перекрестной связи. Примечательно, что эти находки указывают на то, что синтез трансфузии предшествует расцеплению ICL.Эта модель также отличается от более ранних моделей, вызывающих столкновение одной репликационной вилки с ICL (Niedernhofer et al., 2005). На небольшой матрице плазмидной ДНК конвергенция 2 вилок на ICL является правилом. Однако на хромосоме неясно, как часто это явление будет иметь место. Расстояние между источниками репликации колеблется от 150 до 500 т.п.н. в соматических клетках, и, учитывая, что ICL активируют контрольную точку, которая снижает плотность активных источников, разумно полагать, что значительная часть ICL будет удовлетворена одной репликационной вилкой. .Следовательно, эти два режима зависимой от репликации репарации ICL могут сосуществовать.
Используя ДНК-субстрат, содержащий MMC-подобный аддукт ICL, который значительно искажает спираль ДНК, другое исследование обнаружило, что восстановление ICL может происходить посредством репликационно-зависимых и независимых механизмов (Ben-Yehoyada et al., 2009). Ремонт связан со значительным синтезом ДНК (Ben-Yehoyada et al., 2009). Наблюдение за тем, что репарация ICL может происходить в отсутствие репликации ДНК в экстрактах Xenopus и после трансфекции ICL-содержащей плазмиды в G1-арестованные клетки млекопитающих (Ben-Yehoyada et al., 2009) согласуется с накоплением доказательств репарации ICL в G1, обсужденных выше (Hlavin et al., 2010; Muniandy et al., 2010; Sarkar et al., 2006; Shen and Li, 2010). Некоторые из различий, описанных в Xenopus , могут быть связаны с разными типами используемых ICL. Как упоминалось выше, дрожжи используют разные пути репарации для репарации ICL, генерируемых разными сшивающими агентами (Beljanski et al., 2004). Аналогичным образом было показано, что в экстрактах клеток млекопитающих уровень расцепления 5′-GC-3 ‘ICL (искажающий) был в 10 раз выше, чем для 5′-CG-3’ ICL (не искажающий). .Эти результаты демонстрируют, что искажение спирали существенно влияет на распознавание ICL и последующую репарацию (Smeaton et al., 2008).
Разработка сайт-специфичных систем шаблонов ICL открывает большие перспективы для будущих исследований ICL, поскольку эти субстраты предоставляют возможность исследовать процессы ICL, в частности, в отсутствие других форм повреждения ДНК. Использование этих субстратов поможет разграничить молекулярные этапы, участвующие в процессинге ICL, и задействованные белки.Сайт-специфическая природа этих субстратов ICL позволяет определять эффективность восстановления и анализировать промежуточные продукты восстановления и ошибки ремонта. Кроме того, способность вводить сайт-специфические аддукты ICL в клетки млекопитающих позволяет провести in vivo, исследования репарации ICL и передачи сигналов. Это также должно служить прогностическим инструментом для оценки способности клеток больных раком восстанавливать ICL и впоследствии прогнозировать реакцию пациентов на перекрестно связывающие препараты. Аналогичным образом, этот инструмент можно использовать для оценки эффективности терапевтических средств, направленных на восстановление поперечных сшивок.
Белки FA и активация контрольных точек
Путь FA был связан с активностью контрольных точек множеством способов. Сложность этих взаимодействий видна из того факта, что FA участвует выше и ниже передачи сигналов киназы контрольных точек. Исследования в бесклеточных экстрактах Xenopus с использованием ДНК-субстрата, содержащего аддукт 5′-GC-3 ‘ICL, который искажает спираль ДНК, показали, что ICL активирует устойчивые репликационно-зависимые и независимые контрольные точки. После распознавания повреждения ICL путь FA играет важную роль в индукции ATR / Chk1-зависимой контрольной точки (Ben-Yehoyada et al., 2009). Это согласуется с независимой от репликации специфической ассоциацией FA core и ID комплексов с псораленовой ICL (Shen et al., 2009). Истощение корового белка FA (FANCL) или FANCD2 нарушает передачу сигналов контрольной точки. Активность контрольной точки измеряли по способности ICL-содержащей ДНК ингибировать репликацию неповрежденной плазмиды в trans . Ингибирование пути FA также ингибировало фосфорилирование CHK1. В соответствии с этим открытием, истощение белков FA предотвращает рекрутирование RPA и ATR в ICL.Это устанавливает, что путь FA действует выше сигнального пути RPA-ATR-CHK1. Важно отметить, что рекрутирование RPA и ATR в плазмиды, несущие УФ-поражения, не затрагивается ингибированием пути FA, таким образом идентифицируя ICL-специфичную сигнальную функцию для пути FA (Ben-Yehoyada et al., 2009). Генерацию промежуточных звеньев ssDNA-RPA при обнаружении ICL с прогрессирующей вилкой репликации можно представить как следствие остановки вилки и обработки ICL (). Напротив, механизм, ответственный за генерацию оцДНК-RPA в отсутствие репликации ДНК, неизвестен ().В частности, остается определить, является ли рекрутирование белков FA предпосылкой для расцепления ICL и / или требуется ли разрез (и) на ICL для активации контрольной точки.
Регуляция клеточного цикла передачи сигналов от перекрестной сшивки и репарации перекрестных сшивок
A. Каноническая контрольная точка репликации, активируемая повреждением на одной цепи ДНК. Повреждение приводит к остановке ДНК-полимеразы, которая становится отщепленной от ДНК-геликазы MCM. Полученные промежуточные соединения оцДНК-RPA активируют путь контрольной точки ATR-CHK1 (Byun et al., 2005). Эта контрольная точка активируется в ответ на УФ-повреждения, ингибирование ДНК-полимеразы, а также внутрицепочечное сшивание, генерируемое сшивающими агентами, такими как цисплатин.
B. Активация КПП и ремонт ICL в G1. В G1 активируются комплексы ядра и ID, связанные с повреждением ICL и контрольной точкой. Неизвестно, накапливается ли оцДНК-RPA до или как следствие разреза в непосредственной близости от перекрестной сшивки. Представлено несколько гипотетических режимов оцДНК-RPA.Для ясности накопление оцДНК-RPA показано только с одной стороны ICL, но может происходить с обеих сторон. Ремонт будет продолжаться после расцепления через синтез трансформации и NER.
C. Активация контрольной точки и ремонт ICL в фазе S. В S-фазе FANCD1 / FANCJ / FANCN (Downstream FA) ассоциируются с вилкой, остановившейся на ICL, независимо от комплексов ядра и ID. Гипотетически оцДНК-RPA могла быть образована после разрезания и / или расцепления ICL на отстающей цепи.Кроме того, резецированный DSB, созданный после разреза, может быть дополнительным источником оцДНК-RPA для активации контрольной точки. Обратите внимание, что геликаза MCM, как полагают, загружается вокруг дцДНК и поэтому не может раскручивать ДНК, прошедшую ICL. Следовательно, механизм активации контрольной точки в ICL, по-видимому, отличается от разобщения полимеразы-геликазы, описанного в A. Ремонт будет происходить после расцепления посредством синтеза трансформации, NER и HDR. Примечание: цветная версия рисунка доступна в Интернете.
FANCM участвует в ответе контрольных точек, опосредованном ATR, независимо от основного комплекса FA (Collis et al., 2008). В отсутствие FANCM, TOPBP1 не удерживается на хроматине и фосфорилирование нижестоящих мишеней ATR является дефектным (Luke-Glaser et al., 2010; Schwab et al., 2010). Интересно, что FANCM, по-видимому, играет центральную роль в петле обратной связи при активации контрольных точек, поскольку CHK1 предотвращает зависимую от протеасомы деградацию FANCM при повреждении ДНК.
Белки
FA также необходимы после киназ контрольных точек для правильного выполнения передачи сигналов контрольных точек.Это было впервые продемонстрировано, показывая, что фосфорилирование FANCD2 с помощью ATM после IR необходимо для активации контрольной точки S-фазы (Taniguchi et al., 2002b). Сходным образом было показано, что ядерный комплекс FA и FANCD2 регулируют ATR-зависимую контрольную точку внутри S-фазы в ответ на искажение ICL, индуцированное псораленом (Pichierri and Rosselli, 2004a, b). Напротив, другие исследования показали, что путь FA не регулирует контрольную точку внутри S-фазы после лечения MMC, которая генерирует неискажающие ICL (Andreassen et al., 2004). Эти различия могут отражать разные ответы клеток, дефицитных по разным генам FA, и / или разные ответы на разные сшивающие агенты, приводящие к поражениям ICL с разными свойствами ().
Пути контрольных точек также регулируют путь FA и, в частности, активацию FANCD2. ATR необходим для эффективного убиквитилирования FANCD2 (Andreassen et al., 2004). Как ATR, так и BRCA1, как было показано, способствуют убиквитилированию FANCD2 в ответ на повторную репликацию (Zhu and Dutta, 2006b).Путь FA, по-видимому, участвует в петле обратной связи в ответ на репликацию, поскольку основной комплекс FA, но не FANCD2, необходим для активации контрольных точек в реплицирующихся клетках (Zhu and Dutta, 2006a).
Путь FA и репликация ДНК
Путь FA не только активируется после обработки сшивающими агентами, но также реагирует на лечение, которое влияет на репликацию ДНК (например, индукцию репликационного стресса). Путь FA также активируется во время нормальной S-фазы, а комплекс ID связывается с хроматином во время S-фазы в отсутствие экзогенного повреждения ДНК (Mi and Kupfer, 2005; Sobeck et al., 2006). Нарушение пути FA ведет к накоплению DSB в отсутствие экзогенного повреждения (Sobeck et al., 2006), указывая тем самым, что путь FA играет роль в предотвращении накопления повреждений во время невозмущенной репликации ДНК. Природа структуры (структур) ДНК, которые распознаются и обрабатываются путем FA во время репликации ДНК, остается неясной. В бесклеточных экстрактах путь FA, как видно по убиквитилированию FANCD2, активируется различными структурами (Sobeck et al., 2007).
Core комплекс FA взаимодействует с BLM, Topoisomerase IIα и RPA в комплексе BRAFT (Meetei et al., 2003b) и необходим для ICL-зависимого фосфорилирования BLM и рекрутирования в ядерные фокусы (Pichierri et al., 2004). Было показано, что как ATR, так и BLM функционируют на остановившихся вилках репликации, чтобы стабилизировать вилки во время процессов восстановления и предотвратить коллапс вилки репликации. Нарушение пути FA увеличивает экспрессию хрупких сайтов, а также количество разрывов и разрывов в известных хрупких сайтах (Howlett et al., 2005). Белки FANCD2, FANCI и BLM связываются с ультратонкими мостиками (UFB) во время митоза. Считается, что эти структуры отражают неправильную репликацию локусов ломких сайтов (Chan et al., 2009). Более того, FANCJ и FANCM являются ферментами, которые могут обрабатывать структуры ДНК, возникающие в репликационных вилках (Schwab et al., 2010; Sommers et al., 2009; Wu et al., 2008). ATM и ATR могут фосфорилировать ряд белков FA, и многие из этих фосфорилирования необходимы для эффективной активации пути FA во время репликации ДНК (Ho et al., 2006; Мацуока и др., 2007; Qiao et al., 2004; Wang et al., 2007; Ямасита и др., 1998).
FA белки могут специфически связываться в вилке репликации ДНК, где они готовы обнаруживать и реагировать на повреждение ДНК и стабилизировать вилку репликации (Wang et al., 2008). Нарушение пути FA после инактивации FANCL приводит к нарушению перезапуска репликации в коллапсированных репликационных вилках после обработки MMC или камптотецином (CPT), ингибитором топоизомеразы I (Wang et al., 2008). Сходным образом клетки, дефицитные по FANCM, дефектны в возобновлении репликации ДНК после лечения CPT (Luke-Glaser et al., 2010; Schwab et al., 2010). В то время как MMC генерирует ICL (среди других повреждений ДНК), а CPT в конечном итоге приводит к образованию DSB, оба лечения приводят к образованию повреждений ДНК, блокирующих репликационную вилку. Требование пути FA в перезапуске репликации после обеих обработок указывает на то, что путь FA функционирует для стабилизации репликационной вилки, когда он встречает физическое повреждение, блокирующее вилку, и / или для координации повторной сборки репликационной вилки и перезапуска репликации после удаление очага поражения.
Белки ЖК при раке
Путь ЖК во многих отношениях связан с раком. Во-первых, анемия Фанкони связана с предрасположенностью к раку. Во-вторых, поскольку путь FA имеет решающее значение для восприятия и восстановления ICL, его целостность определяет чувствительность или устойчивость к химиотерапевтическим препаратам, которые генерируют ICL.
Взаимосвязь между анемией Фанкони и предрасположенностью к раку хорошо известна. Почти у 25% пациентов с ФА развиваются злокачественные новообразования, особенно миелодиспластический синдром, острый миелоцитарный лейкоз и плоскоклеточный рак головы и шеи (Kutler et al., 2003). Однако совсем недавно белки FA были вовлечены в семейные и спорадические раковые заболевания за пределами популяции пациентов с FA. FANCD1, FANCJ и FANCN вовлечены в предрасположенность к раку молочной железы, поскольку были идентифицированы усекающие мутации в каждом из этих генов (Erkko et al., 2007; Rahman et al., 2007; Reid et al., 2007; Seal et al., 2006; Turnbull, Rahman, 2008; Xia et al., 2007). Мутации FANCD1 также были связаны с раком яичников, простаты, желудка и поджелудочной железы (Friedenson, 2005), а укорочения FANCN / PALB2 были связаны с раком простаты (Erkko et al., 2007). RAD51C, который недавно был вовлечен в FA-подобное расстройство (Vaz et al., 2010), также участвует в предрасположенности к раку груди и яичников (Meindl et al., 2010). Моноаллельные доминантные мутации наблюдались в семьях с раком груди и яичников, напоминающими семьи с дефицитом BRCA2 (Levy-Lahad, 2010; Meindl et al., 2010). Метилирование промотора также было идентифицировано как метод инактивации ЖК. Метилирование промотора FANCF было впервые обнаружено в связи с раком яичников Taniguchi et al.(Taniguchi et al., 2003), и это открытие было подтверждено несколькими другими группами (Dhillon et al., 2004; Wang et al., 2006). Помимо рака яичников, метилирование промотора FANCF связано с острым миелоидным лейкозом (Tischkowitz et al., 2003), множественной миеломой (Chen et al., 2005; Hazlehurst et al., 2003), раком мочевого пузыря (Neveling et al. , 2007), рак шейки матки (Narayan et al., 2004), карциномы головы и шеи и немелкоклеточный рак легких (Marsit et al., 2004). Метилирование промотора других членов комплекса ядра FA, FANCC, FANCG и FANCL, также вовлечено в рак поджелудочной железы (Couch et al., 2005; Роджерс и др., 2004; van der Heijden et al., 2003), рака груди (Sinha et al., 2008) и лейкемии (Hess et al., 2008). Сообщалось о метилировании промотора PALB2 в опухолях молочной железы (Potapova et al., 2008). Кроме того, измененная экспрессия FANCL из-за варианта сплайсинга была обнаружена в клетках рака легких Calu-6 (Zhang et al., 2006), а SNP FANCD2, как было обнаружено, был связан со спорадическим раком груди в испанской популяции (Barroso et al. др., 2006). Генетическая инактивация и эпигенетическое молчание пути FA при спорадических раковых опухолях подчеркивают критическую опухолевую супрессорную функцию этого пути.
Реакция раковых клеток, чувствительность или устойчивость, на препараты, повреждающие ДНК, такие как сшивающий агент цисплатин, может варьироваться в зависимости от состояния определенных путей репарации ДНК. Например, чувствительность к цисплатину в половых клетках семенников и клетках немелкоклеточного рака легкого связана со сниженной экспрессией белков эксцизионной репарации нуклеотидов XPA и ERCC1, соответственно (Fujii et al., 2008; Koberle et al., 1999; Vilmar and Sorensen , 2008; Welsh et al., 2004). Кроме того, вторичные мутации в BRCA1 опосредуют устойчивость к цисплатину в опухолях яичников с мутацией BRCA1 (Swisher et al., 2008). Следовательно, состояние и модуляция путей репарации ДНК могут играть роль в адаптации режимов химиотерапии и повторной сенсибилизации цисплатин-резистентных злокачественных опухолей к лечению, соответственно.
Подобно клеткам пациентов с FA, многие линии раковых клеток и образцы первичных опухолей с нарушением пути FA демонстрируют чувствительность к кросслинкерам. Инактивация пути FA была связана с повышенной чувствительностью к сшивающим агентам, таким как цисплатин, MMC и мелфалан, в яичниках (Taniguchi et al., 2003), поджелудочной железы (Sakai et al., 2008; van der Heijden et al., 2005; van der Heijden et al., 2004), мочевого пузыря (Neveling et al., 2007), множественной миеломы (Yarde et al., 2009) и рака легких (Zhang et al., 2006). Кроме того, дефекты пути FA были связаны с чувствительностью к алкилирующим агентам в клетках глиомы (Chen et al., 2007). Было обнаружено, что дефект пути FA напрямую отвечает за химиочувствительность, поскольку восстановление интактного пути FA отменяет чувствительность и приводит к химиорезистентности (Taniguchi et al., 2003; van der Heijden et al., 2005). Несколько исследований также показали, что целенаправленное нарушение пути FA путем делеции FANCC или FANCG, введение доминантно-отрицательного FANCA, нацеливание siRNA на FANCF или FANCD2 или ингибирование низкомолекулярным ингибитором куркумином напрямую приводит к хемосенсибилизации к цисплатину, мелфалан, алкилирующие агенты и ингибиторы PARP (Chen et al., 2007; Chen et al., 2005; Ferrer et al., 2004; Gallmeier et al., 2006; Sakai et al., 2008). Наконец, инактивация BRCA2 / FANCD1 была связана с чувствительностью к цисплатину при карциноме яичников.Связь между нарушением пути FA и химиочувствительностью определяет модуляцию пути FA как средство хемосенсибилизации. Точно так же выявление дефектов пути FA в опухолях может привести к созданию режимов целевой терапии рака, которые используют преимущества этого опухолеспецифического дефекта.
В то время как нарушение пути FA приводит к чувствительности к цисплатину, реактивация функций пути приводит к развитию устойчивости к цисплатину. Поразительно, что резистентность de novo к BRCA2-дефицитной опухоли яичника, вызванная реверсией точечных мутаций BRCA2, была описана у пациентов и в клетках в культуре (Sakai et al., 2009; Сакаи и др., 2008). Инкубация in vitro чувствительных к цисплатину клеток, содержащих мутацию FANCD1 / BRCA2 или гиперметилирование промотора FANCF, также приводила к приобретенной устойчивости к цисплатину, опосредованной восстановлением пути FA (Sakai et al., 2008; Taniguchi et al., 2003) . В случае метилирования промотора FANCF обработка деметилирующими агентами восстанавливает устойчивость к цисплатину (Taniguchi et al., 2003). Процессы инактивации и реактивации FA проливают свет на механизмы чувствительности и устойчивости к цисплатину, а также предлагают новые методы лечения рака посредством ингибирования пути FA.Первоначально клетка может воспользоваться преимуществом инактивированного пути FA для создания хромосомной нестабильности и ускорения мутагенеза, в течение которого она чувствительна к лечению цисплатином. Однако лечение цисплатином может привести к отбору ревертантных клеток, которые реактивировали путь FA и теперь устойчивы к цисплатину. Следовательно, нацеливание на путь FA может служить для повторной сенсибилизации этих клеток к цисплатину и может использоваться в комбинации с цисплатином для повышения эффективности этого лечения.
Скрининги идентифицировали гены, которые при нарушении регуляции сенсибилизируют клетки FA.Следует отметить, что обе основные ветви путей контрольных точек повреждения ДНК, то есть ATM и ATR-CHK1, сенсибилизируют клетки FA при пониженной регуляции (Chen et al., 2009; Kennedy et al., 2007). Сходным образом была описана небольшая молекула, которая делает клетки FA сверхчувствительными к сшивающим агентам (Gallmeier et al., 2007). Несколько групп также начали поиск ингибиторов пути FA. Chirnomas et al. сообщили о первоначальном обнаружении 4 ингибиторов пути FA / BRCA — вортманнина, H-9, альстерпауллона и куркумина — в результате высокопроизводительного скрининга с использованием человеческих клеток (Chirnomas et al., 2006). Landais et al. разработали анализ на основе бесклеточного экстракта Xenopus на ингибиторы FA (Landais et al., 2009; Landais et al., 2008). Jacquemont и Taniguchi показали, что протеосомные ингибиторы также ингибируют путь FA (Jacquemont and Taniguchi, 2007), а Burkitt и Ljungman показали, что фенилбутират ингибирует путь FA (Burkitt and Ljungman, 2008). Специфическое ингибирование пути FA открывает многообещающее будущее для лечения рака, позволяя нам улучшить эффективность наших текущих схем и преодолеть развитие резистентности.
Заключение
Путь FA играет центральную роль в репарации ICL и поддержании целостности генома. Функционируя выше механизмов передачи сигналов и восстановления контрольных точек, путь FA действует как дозорный при обнаружении ICL. Как только обнаруживается ICL, активируется путь FA и сигнализирует нисходящий поток, чтобы активировать ATR / Chk1-зависимую контрольную точку. Путь FA также выполняет функцию координации белков репарации HDR, TLS и NER при репарации ICL. Во время распознавания и репарации ICL путь FA стабилизирует репликационную вилку, так что репликация может возобновиться после устранения повреждения.В отсутствие интактного пути FA клетки чувствительны к спонтанным и вызванным повреждением ДНК хромосомным разрывам.
Нарушение пути FA было обнаружено при ряде раковых заболеваний и может играть роль в канцерогенезе, внося вклад в нестабильность генома. Инактивация пути FA путем метилирования промотора и усечения мутаций белков FA нарушает целостность генома и способствует чувствительности к цисплатину при многих раковых заболеваниях. Однако резистентность к цисплатину может развиться после начального лечения цисплатином, поскольку путь FA становится реактивированным.Существенная роль пути FA в чувствительности и устойчивости к цисплатину указывает на то, что путь FA является специфической мишенью для лечения рака. Оценка статуса функции пути FA может идентифицировать специфические опухоли, чувствительные к сшивающим агентам. Кроме того, целенаправленное нарушение пути FA может восстановить чувствительность цисплатина к резистентным опухолям, а также может использоваться в качестве комбинированной терапии с различными химиотерапевтическими агентами, включая сшивающие агенты, ионизирующее излучение, алкилирующие агенты и ингибиторы PARP для повышения их эффективности. лечения.
Разработка индивидуализированной химиотерапии и комбинированной терапии со специфическими химиосенсибилизаторами, которые нацелены на пути репарации ДНК, открывают большие перспективы для будущего лечения рака. Однако важно направить эти хемосенсибилизаторы на опухолевые клетки, чтобы избежать нарушения ключевых путей репарации ДНК в соседних нормальных клетках. Кроме того, ингибиторы должны нацеливаться на ключевые нижестоящие белки, чтобы избежать развития компенсирующих механизмов, ведущих к химиорезистентности.Первоначальные открытия низкомолекулярных ингибиторов пути FA и их значение в восстановлении чувствительности к цисплатину идентифицируют путь FA как путь репарации ДНК, который может быть специфически нацелен таким образом. По мере того, как достигается более глубокое понимание ролей и механизмов пути FA и определяются специфические ингибиторы этого пути, целенаправленная модуляция пути FA может играть важную роль во многих химиотерапевтических режимах.
Антиноцицептивные эффекты новых соединений пиразолов, опосредованные каналом ASIC-1α, рецепторами TRPV-1 и μMOR
Пиразолы являются мощными лекарственными каркасами и проявляют широкий спектр биологических активностей, таких как обезболивающее, противовоспалительное и жаропонижающее.В этой статье мы сообщаем об исследованиях, которые мы провели с целью продолжения биологической оценки регио-изомерных соединений пиразола, LQFM-020 (фтор, пара-положение), LQFM-021 (фтор, мета-положение) и LQFM-039. (фтор, орто-положение) в моделях боли, вызванной подкисленным физиологическим раствором, капсаицином и формалином. Мы также исследовали механизмы действия этих соединений с помощью электрофизиологического анализа с использованием метода фиксации напряжения с двумя электродами и гетерологичной экспрессии в ооцитах Xenopus laevis.Это позволило нам изучить различные подтипы калиевых каналов: канал ASIC-1α, рецепторы TRPV-1 и μMOR. Наши результаты показывают, что соединения LQFM-020, LQFM-021 и LQFM-039 (15, 30 или 60 мг / кг -1 ) ингибировали ноцицептивный ответ, индуцированный подкисленным физиологическим раствором, дозозависимым образом. Доза 30 мг / кг -1 ингибировала ноцицептивный ответ, вызванный капсаицином, на 53,3%, 51,4% и 52,1% соответственно. Кроме того, мы обнаружили, что налоксон отменяет антиноцицептивный эффект, вызываемый соединениями в обеих фазах формалинового теста.В электрофизиологических анализах мы наблюдали, что соединения LQFM-020, LQFM-021 и LQFM-039 не модулируют подтипы потенциал-управляемых K + каналов. Напротив, все испытанные соединения ингибировали канал ASIC-1α при pH 4,5 со значениями IC50 96,1, 91,6 и 235,2 мкМ соответственно. Все соединения также ингибировали канал TRPV-1 со значениями IC50 139,1, 212,5 и 159,1 мкМ соответственно. В отличие от мишеней ASIC-1α и TRPV-1, все соединения проявляли агонистическую активность в отношении рецептора μMOR со значением EC50 117.4, 98,9 и 86,3 мкМ соответственно. Таким образом, мы заключаем, что каналы ASIC-1α, TRPV-1 и μMOR являются мишенями, которые непосредственно участвуют в антиноцицептивном эффекте LQFM-020, LQFM-021 и LQFM-039. Кроме того, модификации положений фтора в фенильных аналогах не изменяют анальгетический эффект. Однако LQFM-039 показал более низкое взаимодействие с каналом ASIC-1α.